

Involvement of neurogranin in the modulation of calcium/calmodulin-dependent protein kinase II, synaptic plasticity, and spatial learning: a study with knockout mice
- PMID: 11016969
- PMCID: PMC17183
- DOI: 10.1073/pnas.210184697
Involvement of neurogranin in the modulation of calcium/calmodulin-dependent protein kinase II, synaptic plasticity, and spatial learning: a study with knockout mice
Abstract
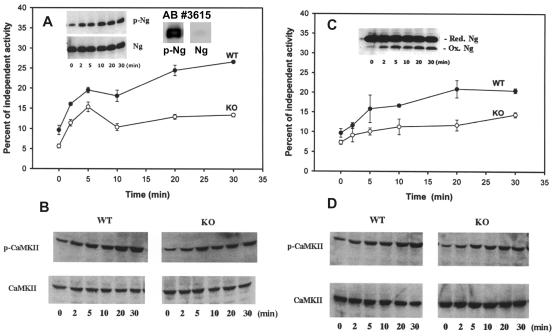
Neurogranin/RC3 is a neural-specific Ca(2+)-sensitive calmodulin (CaM)-binding protein whose CaM-binding affinity is modulated by phosphorylation and oxidation. Here we show that deletion of the Ng gene in mice did not result in obvious developmental or neuroanatomical abnormalities but caused an impairment of spatial learning and changes in hippocampal short- and long-term plasticity (paired-pulse depression, synaptic fatigue, long-term potentiation induction). These deficits were accompanied by a decreased basal level of the activated Ca(2+)/CaM-dependent kinase II (CaMKII) ( approximately 60% of wild type). Furthermore, hippocampal slices of the mutant mice displayed a reduced ability to generate activated CaMKII after stimulation of protein phosphorylation and oxidation by treatments with okadaic acid and sodium nitroprusside, respectively. These results indicate a central role of Ng in the regulation of CaMKII activity with decisive influences on synaptic plasticity and spatial learning.
Figures
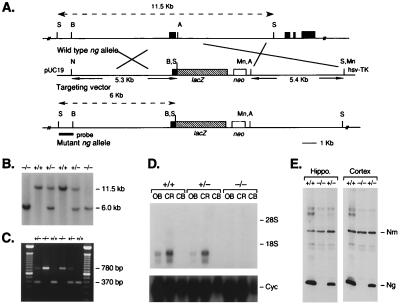
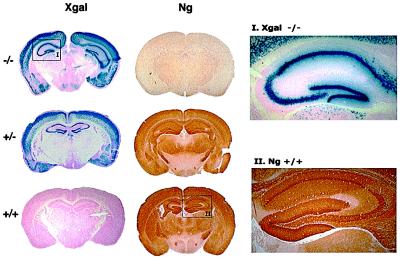
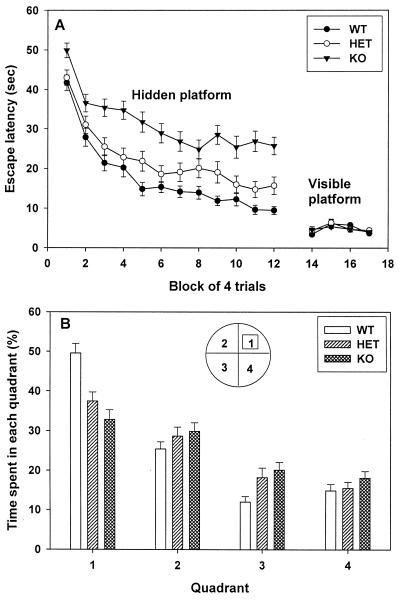
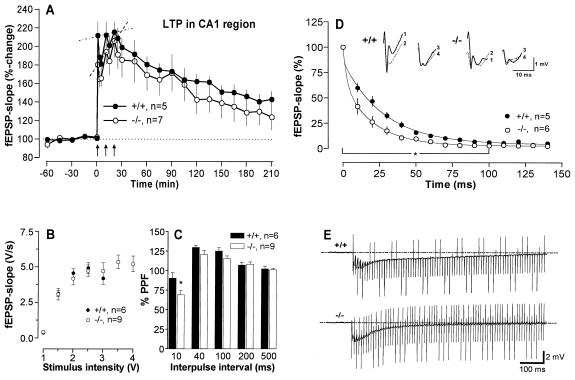
References
-
- Gerendasy D D, Sutcliffe J G. Mol Neurobiol. 1997;15:131–163. - PubMed
-
- Prichard L, Deloulme J C, Storm D R. J Biol Chem. 1999;274:7689–7694. - PubMed
-
- Chakravarthy B, Morley P, Whitfield J. Trends Neurosci. 1999;22:12–16. - PubMed
-
- Baudier J, Deloulme J C, VanDorsselaer A V, Black D, Matthes H W D. J Biol Chem. 1991;266:229–237. - PubMed
-
- Huang K-P, Huang F L, Chen H-C. Arch Biochem Biophys. 1993;305:570–580. - PubMed
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
