

Global analysis of herpes simplex virus type 1 transcription using an oligonucleotide-based DNA microarray
- PMID: 11024119
- PMCID: PMC102029
- DOI: 10.1128/jvi.74.21.9916-9927.2000
Global analysis of herpes simplex virus type 1 transcription using an oligonucleotide-based DNA microarray
Abstract
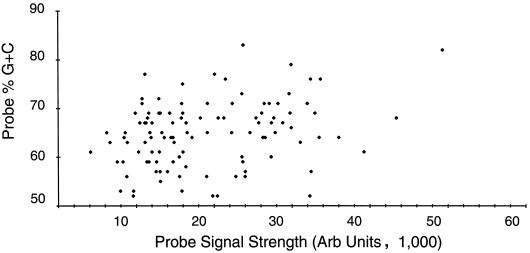
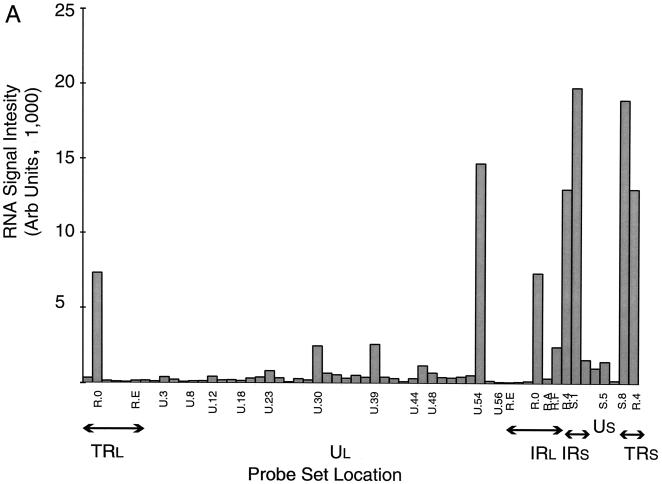
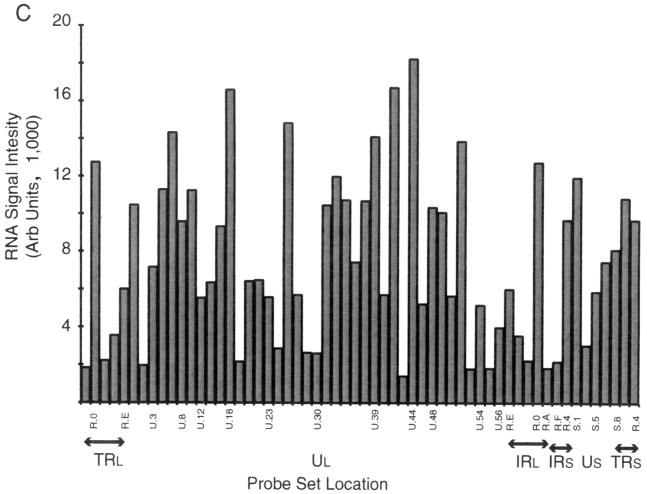
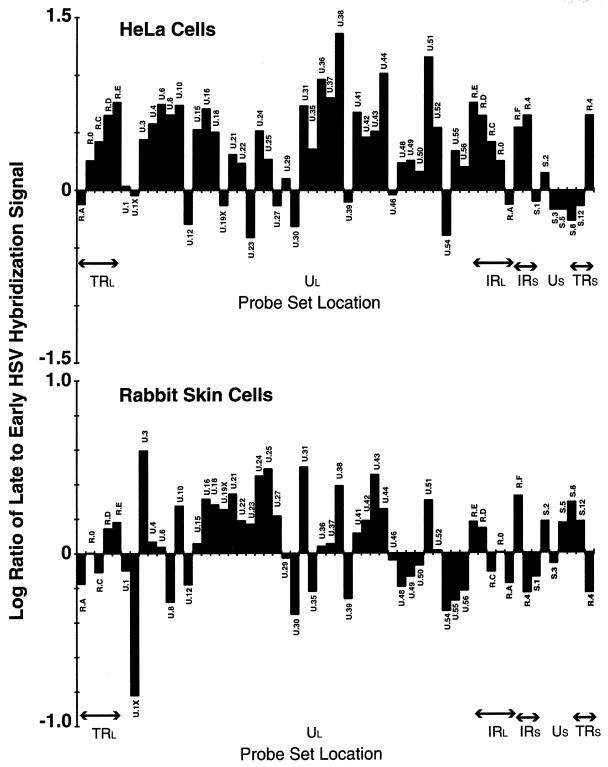
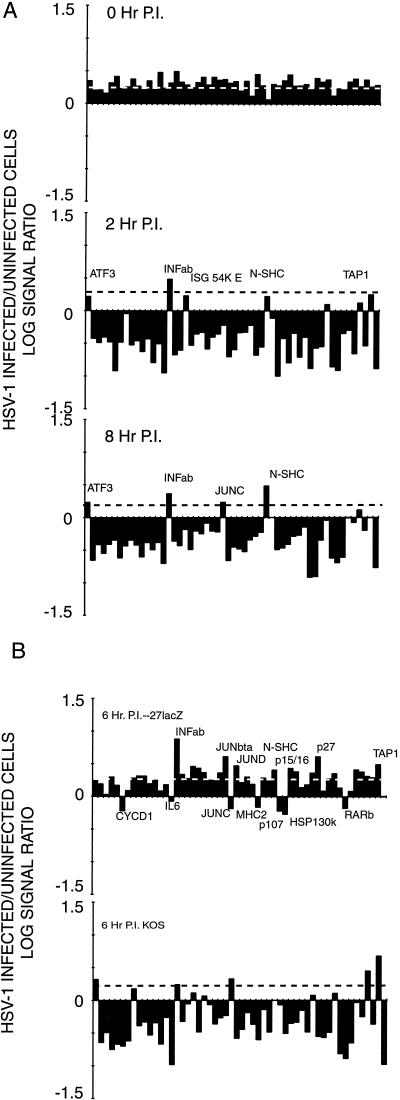
More than 100 transcripts of various abundances and kinetic classes are expressed during phases of productive and latent infections by herpes simplex virus (HSV) type 1. To carry out rapid global analysis of variations in such patterns as a function of perturbation of viral regulatory genes and cell differentiation, we have made DNA microchips containing sets of 75-mer oligonucleotides specific for individual viral transcripts. About half of these are unique for single transcripts, while others function for overlapping ones. We have also included probes for 57 human genes known to be involved in some aspect of stress response. The chips efficiently detect all viral transcripts, and analysis of those abundant under various conditions of infection demonstrates excellent correlation with known kinetics of mRNA accumulation. Further, quantitative sensitivity is high. We have further applied global analysis of transcription to an investigation of mRNA populations in cells infected with a mutant virus in which the essential immediate-early alpha27 (U(L)54) gene has been functionally deleted. Transcripts expressed at 6 h following infection with this mutant can be classified into three groups: those whose abundance is augmented (mainly immediate-early transcripts) or unaltered, those whose abundance is somewhat reduced, and those where there is a significant reduction in transcript levels. These do not conform to any particular kinetic class. Interestingly, levels of many cellular transcripts surveyed are increased. The high proportion of such transcripts suggests that the alpha27 gene plays a major role in the early decline in cellular gene expression so characteristic of HSV infection.
Figures

References
-
- Ben-Hur T, Moyal M, Rosen-Wolff A, Darai G, Becker Y. Characterization of RNA transcripts from herpes simplex virus-1 DNA fragment BamHI-B. Virology. 1989;169:1–8. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
