

Characterization of hepatitis C virus (HCV) and HCV E2 interactions with CD81 and the low-density lipoprotein receptor
- PMID: 11024134
- PMCID: PMC102044
- DOI: 10.1128/jvi.74.21.10055-10062.2000
Characterization of hepatitis C virus (HCV) and HCV E2 interactions with CD81 and the low-density lipoprotein receptor
Abstract
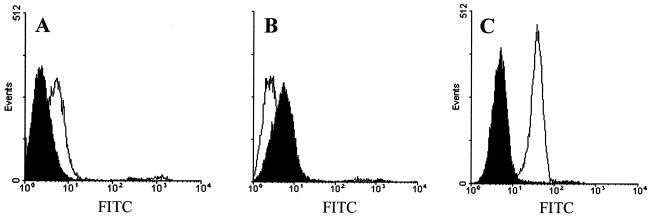
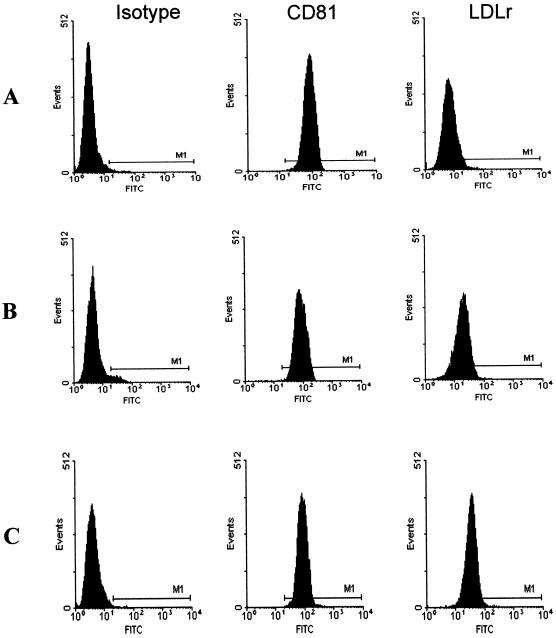
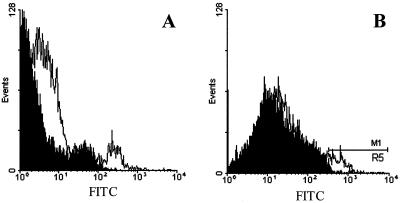
Hepatitis C virus (HCV) or HCV-low-density lipoprotein (LDL) complexes interact with the LDL receptor (LDLr) and the HCV envelope glycoprotein E2 interacts with CD81 in vitro. However, E2 interactions with LDLr and HCV interactions with CD81 have not been clearly described. Using sucrose gradient-purified low-density particles (1.03 to 1.07 g/cm(3)), intermediate-density particles (1. 12 to 1.18 g/cm(3)), recombinant E2 protein, or control proteins, we assessed binding to MOLT-4 cells, foreskin fibroblasts, or LDLr-deficient foreskin fibroblasts at 4 degrees C by flow cytometry and confocal microscopy. Viral entry was determined by measuring the coentry of alpha-sarcin, a protein synthesis inhibitor. We found that low-density HCV particles, but not intermediate-density HCV or controls bound to MOLT-4 cells and fibroblasts expressing the LDLr. Binding correlated with the extent of cellular LDLr expression and was inhibited by LDL but not by soluble CD81. In contrast, E2 binding was independent of LDLr expression and was inhibited by human soluble CD81 but not mouse soluble CD81 or LDL. Based on confocal microscopy, we found that low-density HCV particles and LDL colocalized on the cell surface. The addition of low-density HCV but not intermediate-density HCV particles to MOLT-4 cells allowed coentry of alpha-sarcin, indicating viral entry. The amount of viral entry also correlated with LDLr expression and was independent of the CD81 expression. Using a solid-phase immunoassay, recombinant E2 protein did not interact with LDL. Our data indicate that E2 binds CD81; however, virus particles utilize LDLr for binding and entry. The specific mechanism by which HCV particles interact with LDL or the LDLr remains unclear.
Figures



References
-
- Agnello V, Chung R T, Kaplan L M. A role for hepatitis C virus infection in type II cryoglobulinemia. N Engl J Med. 1992;327:1490–1495. - PubMed
-
- Bouffard P, Hayashi P, Acevedo R, Levy N, Zeldis J. Hepatitis C virus is detected in a monocyte/macrophage subpopulation of peripheral blood mononuclear cells of infected patients. J Infect Dis. 1992;166:1276–1280. - PubMed
-
- Bradley D W, McCaustland K, Cook E H. Posttransfusion non-A, non-B hepatitis in chimpanzees: physicochemical evidence that the tubule forming agent is a small, enveloped virus. Gastroenterology. 1985;88:773–779. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
