

Interaction of the rabies virus P protein with the LC8 dynein light chain
- PMID: 11024151
- PMCID: PMC102061
- DOI: 10.1128/jvi.74.21.10212-10216.2000
Interaction of the rabies virus P protein with the LC8 dynein light chain
Abstract
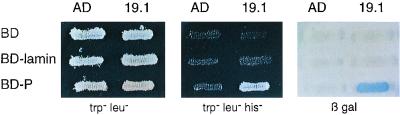
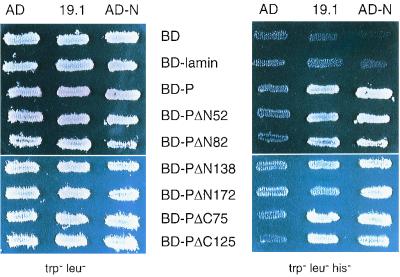
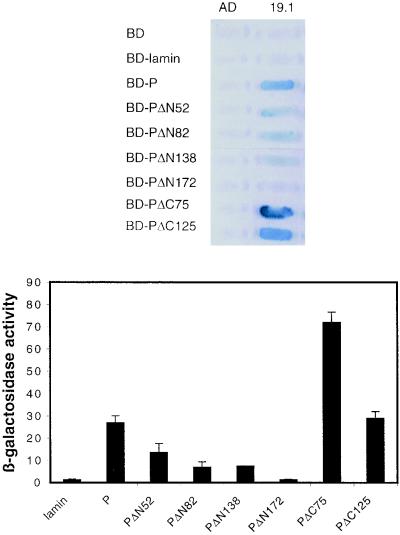
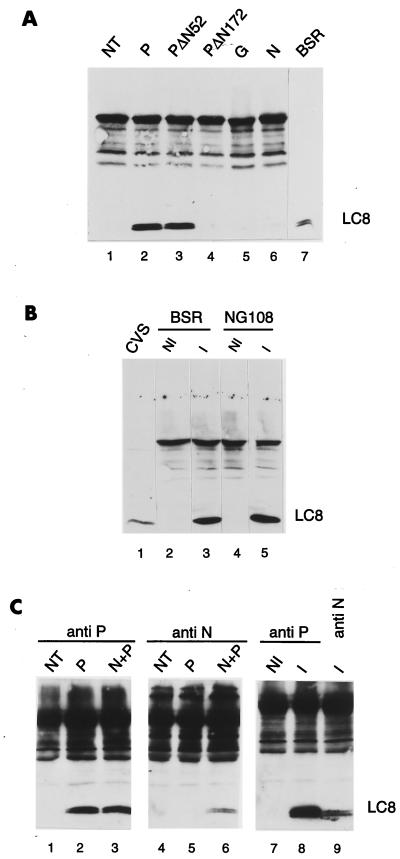
The rabies virus P protein is involved in viral transcription and replication but its precise function is not clear. We investigated the role of P (CVS strain) by searching for cellular partners by using a two-hybrid screening of a PC12 cDNA library. We isolated a cDNA encoding a 10-kDa dynein light chain (LC8). LC8 is a component of cytoplasmic dynein involved in the minus end-directed movement of organelles along microtubules. We confirmed that this molecule interacts with P by coimmunoprecipitation in infected cells and in cells transfected with a plasmid encoding P protein. LC8 was also detected in virus particles. Series of deletions from the N- and C-terminal ends of P protein were used to map the LC8-binding domain to the central part of P (residues 138 to 172). These results are relevant to speculate that dynein may be involved in the axonal transport of rabies virus along microtubules through neuron cells.
Figures

References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
