

doi: 10.1038/35036309.
Temporally coordinated assembly and disassembly of replication factories in the absence of DNA synthesis
Affiliations
- PMID: 11025658
- PMCID: PMC1255923
- DOI: 10.1038/35036309
Item in Clipboard
Temporally coordinated assembly and disassembly of replication factories in the absence of DNA synthesis
Nat Cell Biol.
2000 Oct.
Abstract
Here we show that exposure of aphidicolin-arrested Chinese hamster ovary (CHO) cells to the protein-kinase inhibitors 2-aminopurine or caffeine results in initiation of replication at successively later-replicating chromosomal domains, loss of the capacity to synthesize DNA at earlier-replicating sites, release of Mcm2 proteins from chromatin, and redistribution of PCNA and RPA from early- to late-replicating domains in the absence of detectable elongation of replication forks. These results provide evidence that, under conditions of replicational stress, checkpoint controls not only prevent further initiation but may also be required to actively maintain the integrity of stalled replication complexes.
Figures
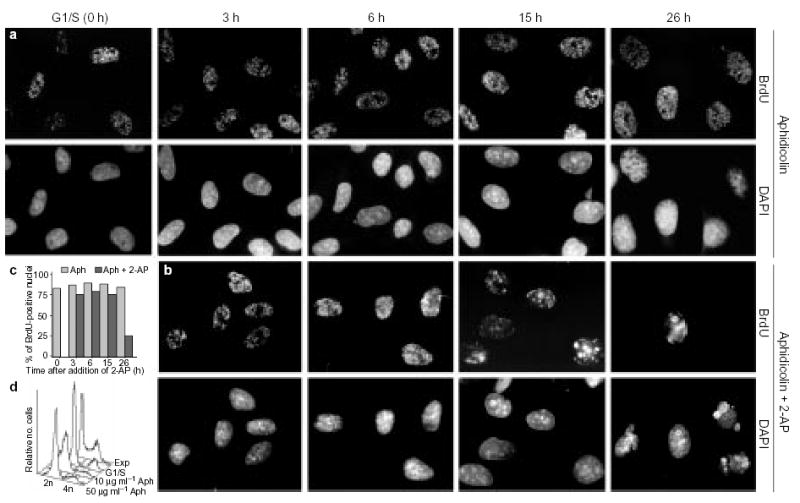
a, b, CHOC 400 metaphase cells were released in fresh medium and arrested at the G1/S border (0 h) in the presence of 50 μg ml−1 aphidicolin (Aph). At this point, 2-AP was added to half of the cell cultures (b), whereas the other half (a) was maintained in aphidicolin only. At the indicated time points, aliquots of cells were washed free of the drugs, pulse-labelled for 10 min with BrdU and stained for BrdU (see Methods). DNA was stained with 4′,6-diamidino-2-phenylindole (DAPI). c, Histogram showing the percentage of BrdU-positive nuclei in a (hatched bars) and b (black bars). d, Flow-cytometry analysis of the DNA content of asynchronously growing cells (Exp), cells arrested at the G1/S boundary, and cells incubated for a further 26 h in the presence of 2-AP and either 10 μg ml−1 or 50 μg ml−1 aphidicolin. As the results shown in a and b were indistinguishable at these two aphidicolin concentrations, the higher concentration was used for all further experiments to prevent any detectable DNA synthesis.
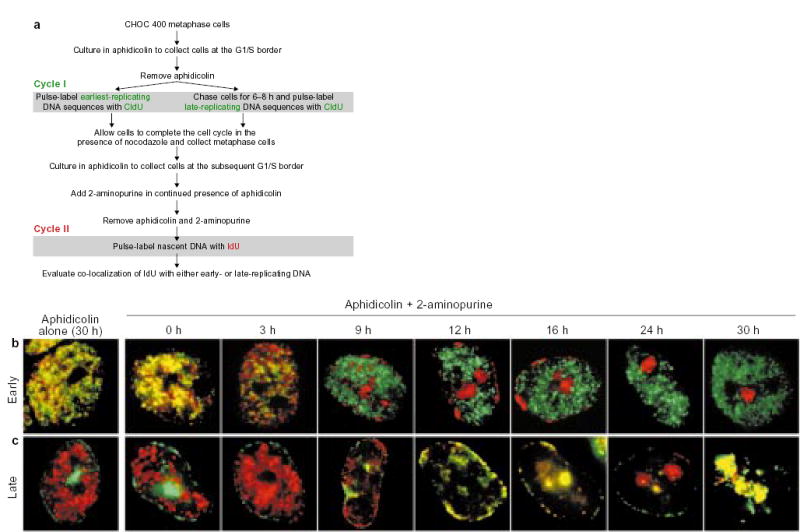
a, Summary of the protocol. b, c, Parallel cultures of CHOC 400 cells were prelabelled with CldU in cycle I and synchronized at the G1/S border of cycle II as outlined in a. Each culture was divided into two and 2-AP was added to one half (Aphidicolin + 2-aminopurine), whereas the other half was maintained in aphidicolin alone. At the indicated times, aliquots of cells were washed free of the drugs, labelled for 10 min with IdU, and then fixed and stained with anti-CldU (green) and anti-IdU (red) antibodies as described. Images show typical nuclei observed at the indicated time points with cells that were prelabelled either early (b) or late (c) in the S phase of cycle I. As the cultures maintained in aphidicolin alone showed co-localization of IdU with the earliest-replicating sequences throughout the entire duration of the experiment (see Fig. 3c, black squares), only the 30-h time point is shown. Similar results were obtained in five independent experiments.
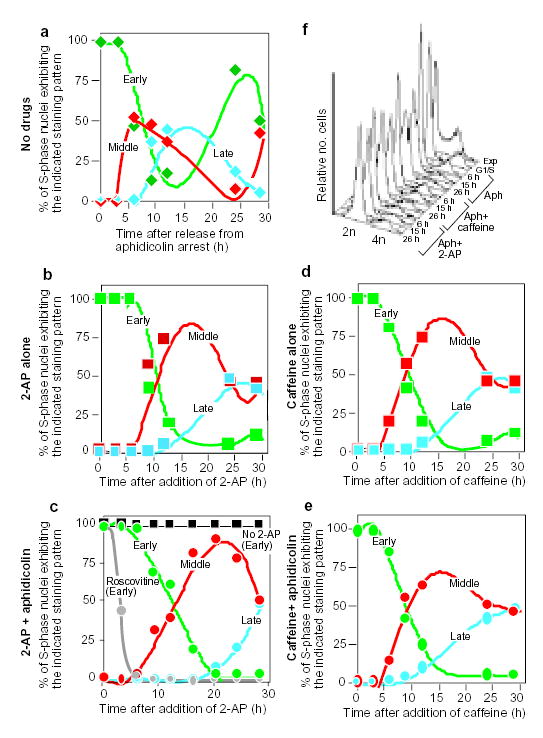
CHOC 400 cells were prelabelled with CldU in cycle I, synchronized at the G1/S border of cycle II as outlined in Fig. 2a, and then divided into aliquots that were treated in parallel as follows. a, Cells were washed free of aphidicolin and allowed to proceed into S-phase in the complete absence of drugs. b, d, Cells were washed free of aphidicolin and allowed to proceed into S phase in the presence of 2-AP (b) or caffeine (d) alone. c, e, Cells were maintained in the continuous presence of aphidicolin in combination with 2-AP (c) or caffeine (e). At the indicated times, cells were washed free of drugs, labelled for 10 min with IdU, and then fixed and stained with anti-CldU and anti-IdU antibodies as in Fig. 2. Percentages of S-phase (IdU-positive) nuclei showing co-localization with either early (type I/II, green), middle (type III, red) or late (type IV and V, blue) CldU-staining patterns were scored at each time point. As controls, aliquots of cells were maintained in aphidicolin alone (c, black squares), or in combination with roscovitine (c, grey circles). Cells maintained in aphidicolin alone showed the same labelling index and exclusively early replication patterns at all time points (see also Fig. 2b, c, aphidicolin alone). When roscovitine was used together with aphidicolin, only the earliest replication patterns were observed and the percentage of IdU-positive nuclei declined with time. f, Aliquots of the cells from a–e were collected at the indicated times and DNA content was measured by flow cytometry. Results were obtained from exponentially growing cells (Exp), cells at 0 h (G1/S), and cells from a (Aph), c (Aph + 2-AP) and e (Aph + caffeine) at the indicated time points. Similar results were obtained in three independent experiments.
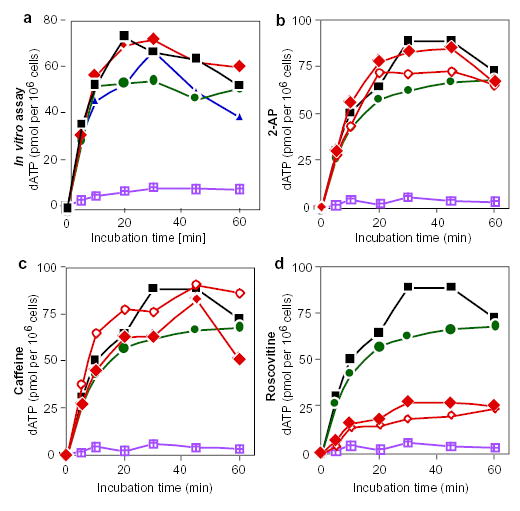
a, Replication run-on assay with drug treatment in vitro. Intact nuclei were prepared from an asynchronous culture of cells and incubated in a replication cocktail supplemented with [α-P]-dATP in the presence of 2-AP (diamonds), caffeine (circles) or roscovitine (triangles). At the indicated time points, aliquots were removed and the amount of acid-precipitable [α-P]-dATP was determined. Only extension of pre-existing replication forks occurs under these conditions. Black squares represent control samples without drugs; violet-hatched squares represent nuclei incubated in vitro in the presence of aphidicolin. Similar results were obtained in three independent experiments. b–d, Replication run-on assays after treatment with 2-AP (b), caffeine (c) or roscovitine (d) in vivo. Asynchronous cultures were incubated for 6 h in the presence of aphidicolin alone (circles), the indicate drug alone (filled diamonds), or the indicated drug in combination with aphidicolin (open diamonds). Intact nuclei were prepared from each cell culture and DNA synthesis was determined as in a. Black squares represent untreated control cultures; violet-hatched squares are reactions carried out in the presence of aphidicolin in vitro.
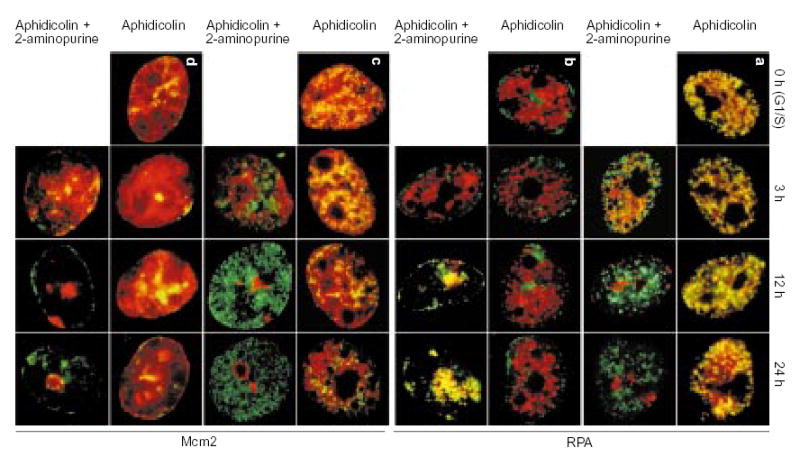
Aliquots of cell cultures, pre-abelled with CldU as in Fig. 2a, were synchronized at the G1/S border of cycle II and maintained in the continuous presence of aphidicolin with or without addition of 2-AP. At the indicated times, aliquots of cells were incubated for 10 min in free medium to match the pulse-labelling time in Fig. 2b, c, fixed and double-stained for CldU (green) and either RPA (a, b; red) or Mcm2 (c, d; red). The distribution of earliest-replicating, CldU-prelabelled chromosomal domains is shown in a and c; the distribution of of mid/late-replicating, CldU-prelabelled chromosomal domains is shown in b and d.

a–c, Cells synchronized at the G1/S border were either released in drug-free medium (a), or maintained in the continuous presence of aphidicolin alone (b) or aphidicolin in combination with 2-AP (c). Nuclei were prepared from these cells at the indicated times and nascent DNA was briefly labelled with [α-P]-dATP in a replication cocktail as described,. As a control, nuclei from exponentially growing cells (Exp) were similarly labelled. Radiolabelled DNA was isolated, sheared and hybridized to a panel of probes representing sequences in the CHOC 400 genome that replicate either early (DHFR), or late (LINES C1 and C3) in S phase. d, e, The relative amounts of hybridization to DHFR probe R (squares), as well as to the C1 (circles) and C3 (diamonds) probes at each time point in a (d) and c (e) were calculated and plotted (see Methods). f–i, Relative amounts of hybridization to all 17 DHFR probes, for the 3-h and 15-h time points in b and c were plotted against the map position of each probe. The horizontal axis includes a diagram of the genomic region encompassed by these probes, including the positions of the DHFR and 2BE2121 loci. Vertical shaded line shows the positions of probes B–R, which encompass the region of peak initiation activity.
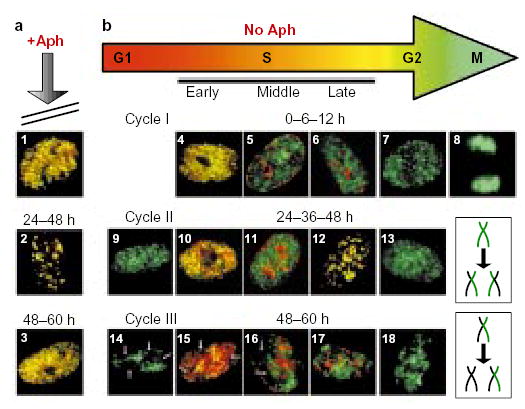
Two asynchronous cultures of AT8052 cells were pulse-labelled for 10 min with CldU, aphidicolin (Aph) was added to one of them (a), and both cultures were grown for a further 60 h. At various time points after CldU labelling (0, 6, 12, 24, 36, 48 and 60 h), aliquots of cells from each culture were washed free of drugs, pulse-labelled with IdU, and then fixed and stained with anti-CldU (green) and anti-IdU (red) antibodies as in Fig. 2. In the presence of aphidicolin, cells were arrested in S phase and the IdU label co-localized with the CldU label in nuclei at all stages of S phase (panels 1 and 3 show examples of early-S-phase nuclei; panel 2 shows a late-S-phase nucleus) throughout the entire chase period. As AT8052 is a primary cell line that could potentially enter into senescence, it was important to verify that cells would have continued to cycle during the chase period. In the absence of aphidicolin (b), the control cell culture grew normally. The size of the nuclei and the type of the IdU replication pattern were used as criteria to evaluate the progression of cells through the cell cycle. During cycle I, the CldU and IdU labelling patterns co-localized at 0 h (panel 4), but had become separated by 6 h (panels 5 and 6) and, at 12 h, large, IdU-negative G2 nuclei (panel 7) and mitotic figures (panel 8) were observed. The appearance of small, CldU-labelled, IdU-negative daughter nuclei (panel 9), often positioned adjacent to each other, indicated progression of cells into the G1 phase of cycle II by 24 h. Later, at 24–48 h, double-labelled S-phase nuclei (panels 10–12) and large, CldU-labelled G2 nuclei (panel 13) re-appeared as the cells moved through cycle II. The appearance of small, IdU-negative nuclei with gaps (arrows) in the CldU labelling pattern (panel 14) indicated that, by 48 h, cells had progressed into cycle III. Gaps correspond to the territories of unlabelled chromosomes,, which were generated after two cell divisions (the mechanism responsible for generation of unlabelled chromosome territories is schematically outlined in the two lower-right boxes). Nuclei with unlabelled chromosome territories were also found to have entered S phase (panels 15–17) and G2 phase (panel 18) of cycle III by the 60-h time point.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
