

Facial visceral motor neurons display specific rhombomere origin and axon pathfinding behavior in the chick
- PMID: 11027227
- PMCID: PMC6772854
- DOI: 10.1523/JNEUROSCI.20-20-07664.2000
Facial visceral motor neurons display specific rhombomere origin and axon pathfinding behavior in the chick
Abstract
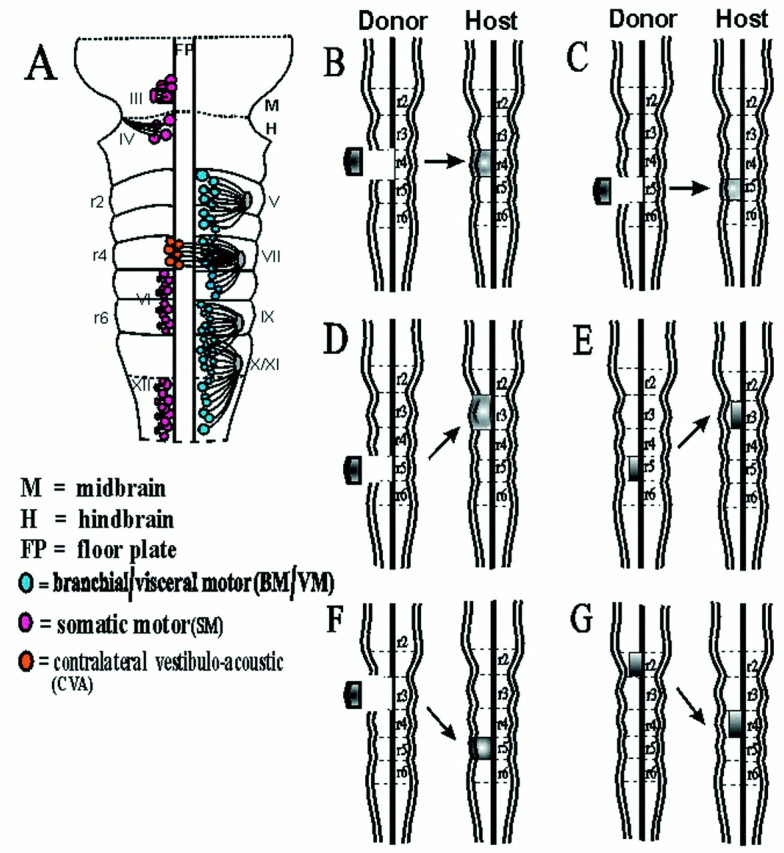
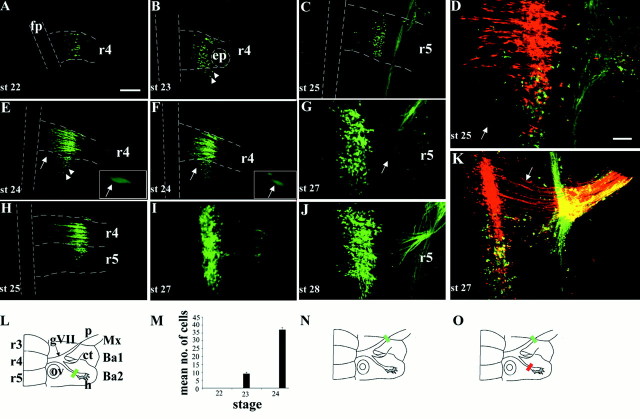
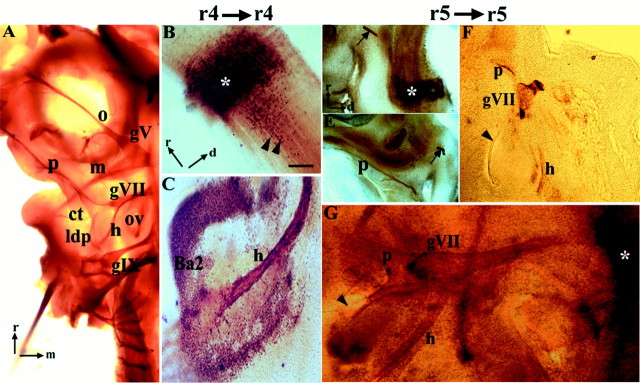
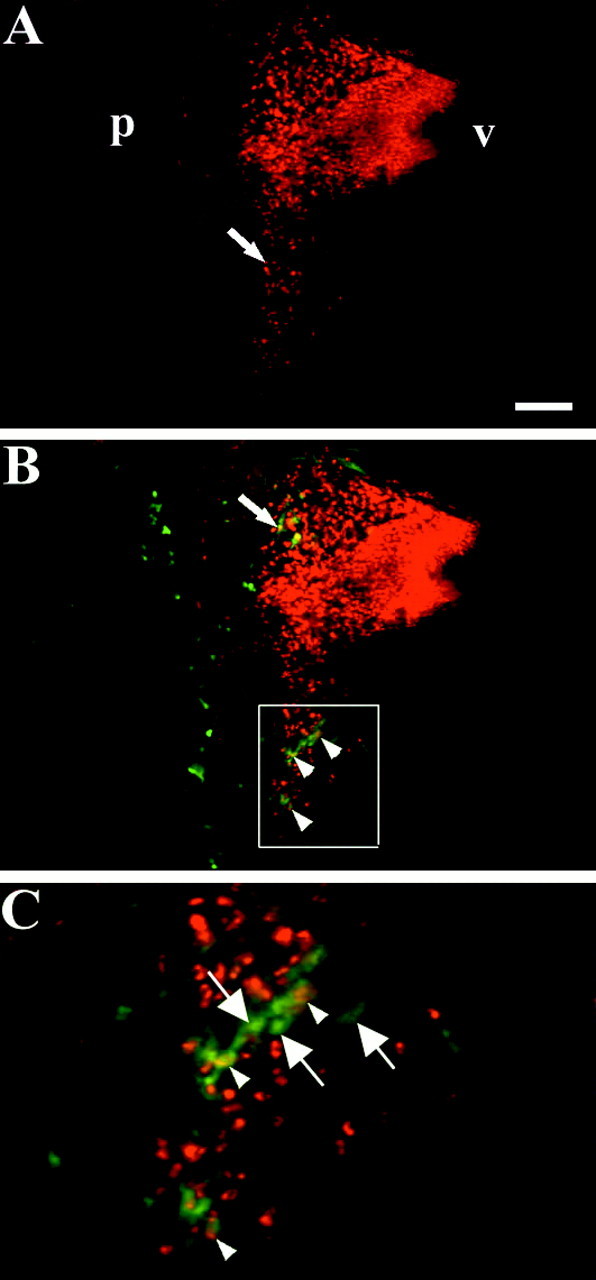
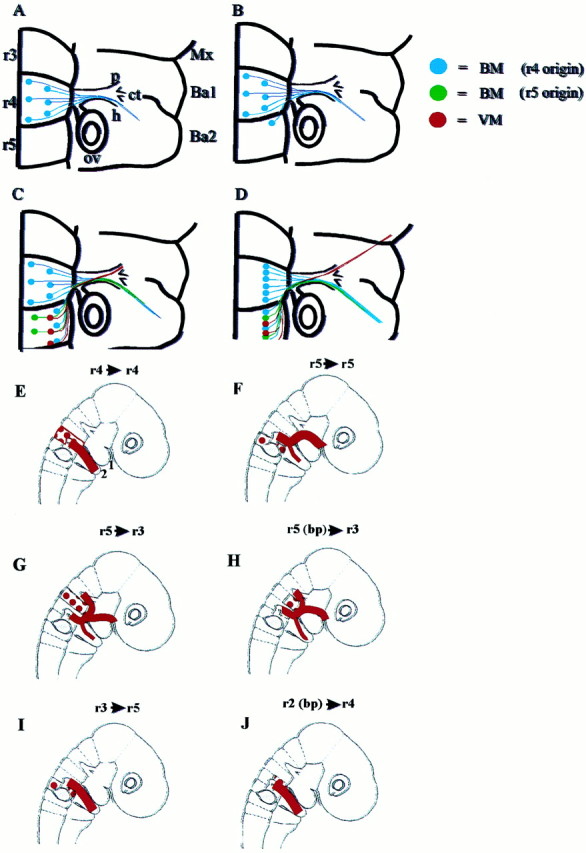
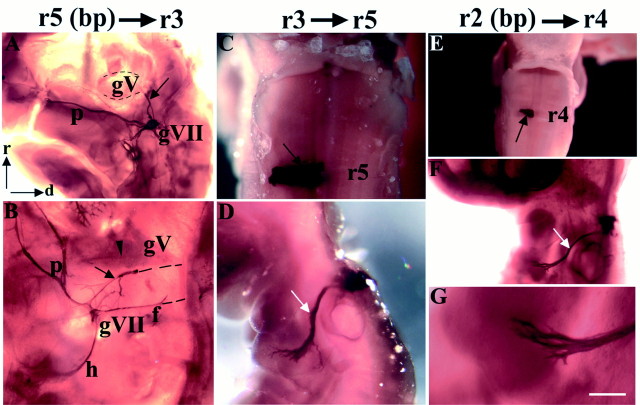
In the chick embryo, facial motor neurons comprise branchiomotor and visceral motor subpopulations, which innervate branchial muscles and parasympathetic ganglia, respectively. Although facial motor neurons are known to develop within hindbrain rhombomere 4 (r4) and r5, the precise origins of branchiomotor and visceral motor neuron subpopulations are unclear. We investigated the organization and axon pathfinding of these motor neurons using axonal tracing and rhombomere transplantation in quail-chick chimeras. Our results show that a large majority of branchiomotor neurons originate in r4 but that a cohort of these neurons undergoes a caudal migration from r4 into r5. By contrast, visceral motor neurons develop exclusively in r5. We found that a striking property of facial visceral motor neurons is the ability of their axons to navigate back to appropriate ganglionic targets in the periphery after heterotopic transplantation. These results complement previous studies in which heterotopic facial branchiomotor neurons sent axons to their correct, branchial arch, target. By contrast, when trigeminal branchiomotor neurons were transplanted heterotopically, we found that they were unable to pathfind correctly, and instead projected to an inappropriate target region. Thus, facial and trigeminal motor neuron populations have different axon pathfinding characteristics.
Figures
References
-
- Auclair F, Valdes N, Marchand R. Rhombomere-specific origin of branchial and visceral motoneurones of the facial nerve in the rat embryo. J Comp Neurol. 1996;369:451–461. - PubMed
-
- Bell E, Wingate RJ, Lumsden A. Homeotic transformation of rhombomere identity after localized Hoxb1 misexpression. Science. 1999;284:2168–2171. - PubMed
-
- Caton A, Hacker A, Naeem A, Livet J, Maina F, Klein R, Birchmeier C, Guthrie S. The branchial arches and HGF are growth-promoting and chemoattractant for cranial motor axons. Development. 2000;127:1751–1766. - PubMed
-
- D'Amico-Martel A, Norden DM. Contributions of placodal and neural crest cells to avian cranial peripheral ganglia. Am J Anat. 1983;166:445–468. - PubMed
-
- Fritzsch B, Nichols DH. DiI reveals a prenatal arrival of efferents at the differentiating otocyst of mice. Hear Res. 1993;65:51–60. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources