

Spatial generalization from learning dynamics of reaching movements
- PMID: 11027245
- PMCID: PMC6772893
- DOI: 10.1523/JNEUROSCI.20-20-07807.2000
Spatial generalization from learning dynamics of reaching movements
Abstract
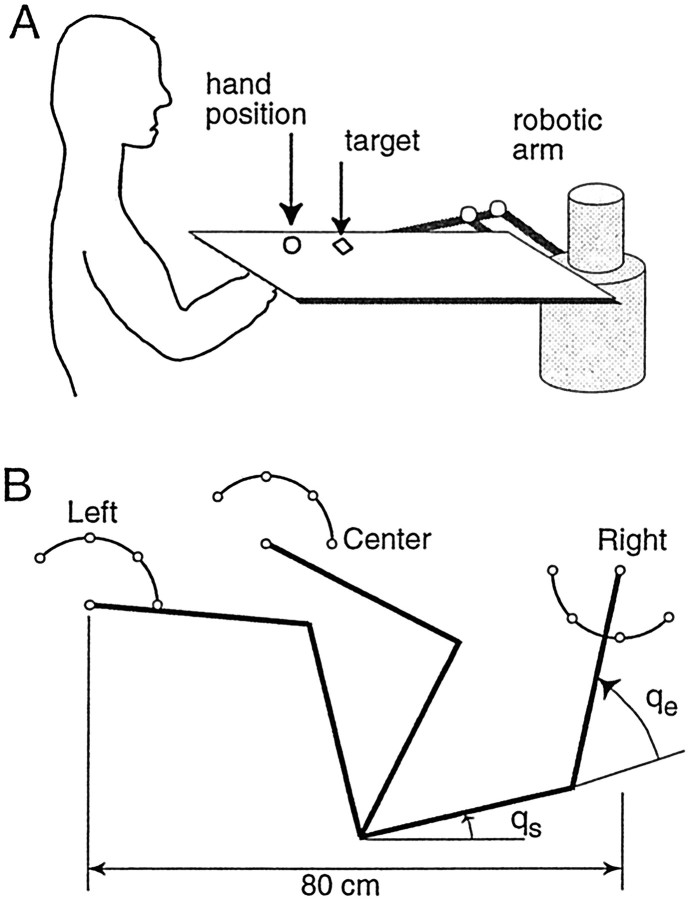
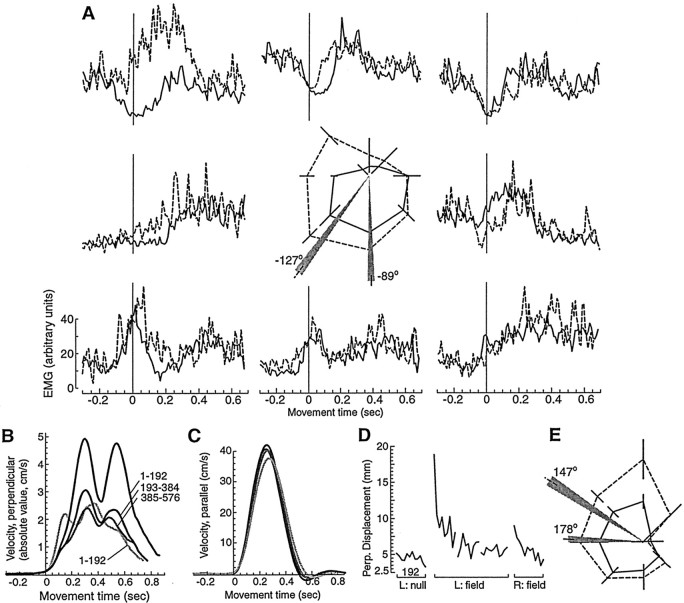
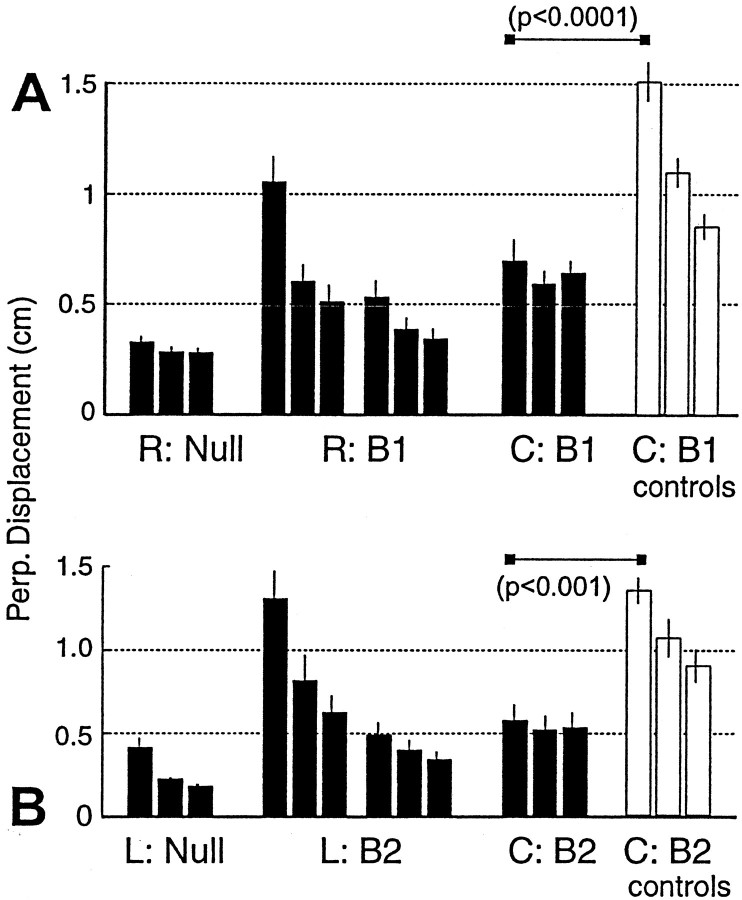
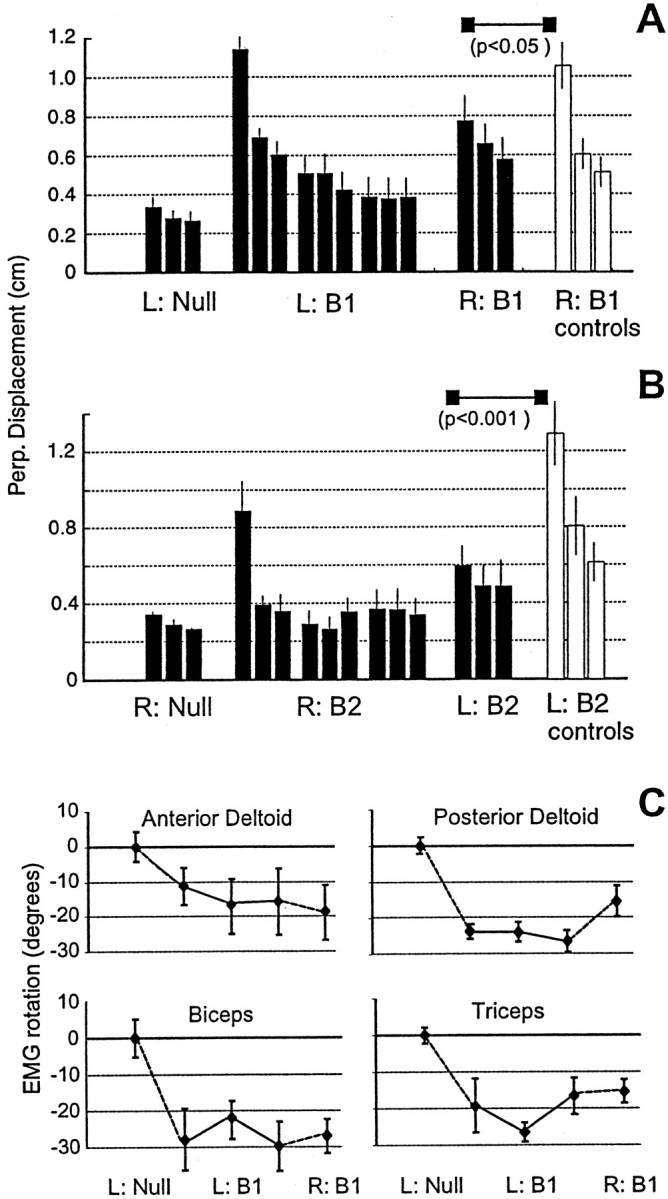
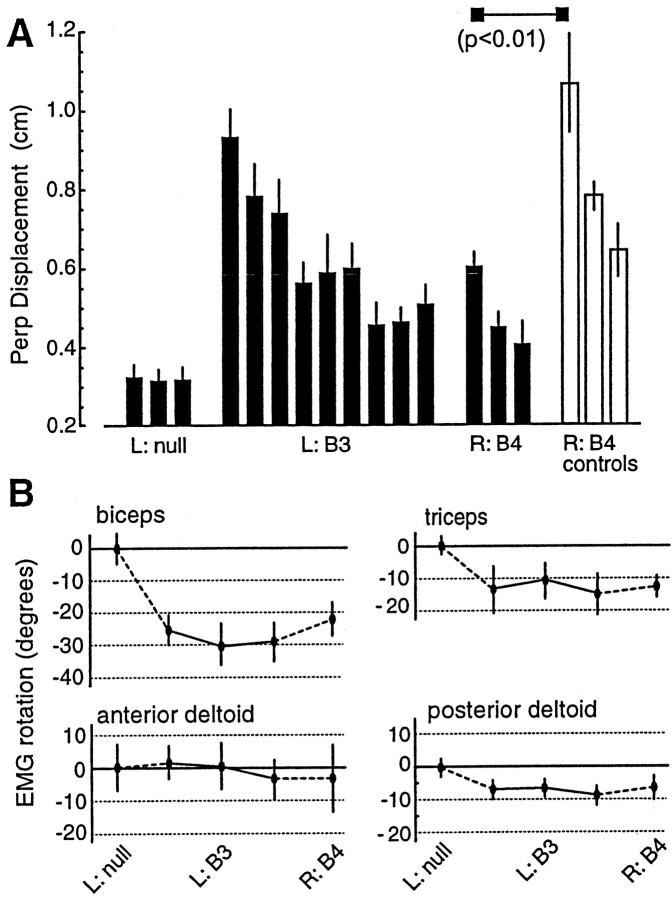
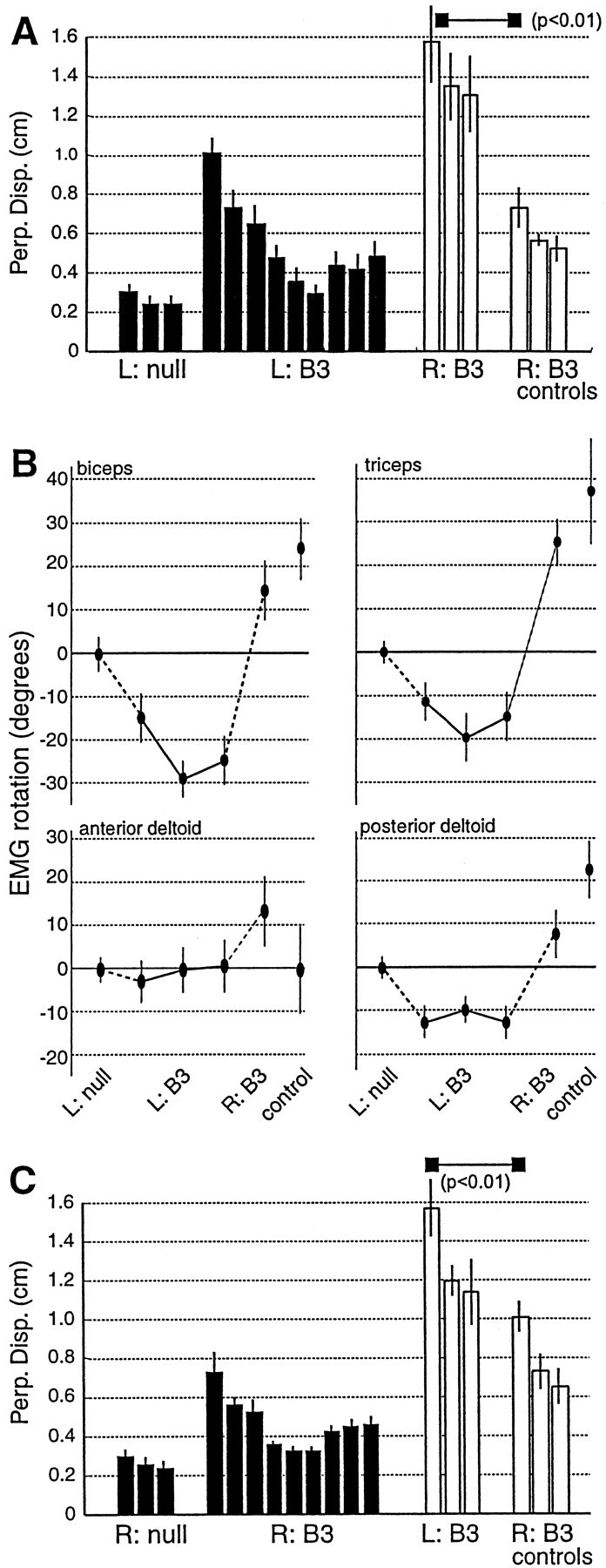
When subjects practice reaching movements in a force field, they learn a new sensorimotor map that associates desired trajectories to motor commands. The map is formed in the brain with elements that allow for generalization beyond the region of training. We quantified spatial generalization properties of these elements by training in one extreme of the reachable space and testing near another. Training resulted in rotations in the preferred direction (PD) of activation of some arm muscles. We designed force fields that maintained a constant rotation in muscle PDs as the shoulder joint rotated in the horizontal plane. In such fields, training in a small region resulted in generalization to near and far work spaces (80 cm). In one such field, the forces on the hand reversed directions for a given hand velocity with respect to the location of original training. Despite this, there was generalization. However, if the field was such that the change in the muscle PDs reversed as the work spaces changed, then performance was worse than performance of naive subjects. We suggest that the sensorimotor map of arm dynamics is represented in the brain by elements that globally encode the position of the arm but locally encode its velocity. The elements have preferred directions of movement but are modulated globally by the position of the shoulder joint. We suggest that tuning properties of cells in the motor system influence behavior and that this influence is reflected in the way that we learn dynamics of reaching movements.
Figures
References
-
- Ahissar M, Hochstein S. Task difficulty and the specificity of perceptual learning. Nature. 1997;387:401–405. - PubMed
-
- Benda BJ, Gandolfo F, Li CSR, Tresch MC, DiLorenzo D, Bizzi E. Neuronal activities in M1 of a macaque monkey during reaching movements in a viscous force field. Soc Neurosci Abstr. 1997;23:1556.
-
- Brashers-Krug T, Shadmehr R, Bizzi E. Consolidation in human motor memory. Nature. 1996;382:252–255. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous