

In yeast, the 3' untranslated region or the presequence of ATM1 is required for the exclusive localization of its mRNA to the vicinity of mitochondria
- PMID: 11027259
- PMCID: PMC86399
- DOI: 10.1128/MCB.20.21.7881-7892.2000
In yeast, the 3' untranslated region or the presequence of ATM1 is required for the exclusive localization of its mRNA to the vicinity of mitochondria
Abstract
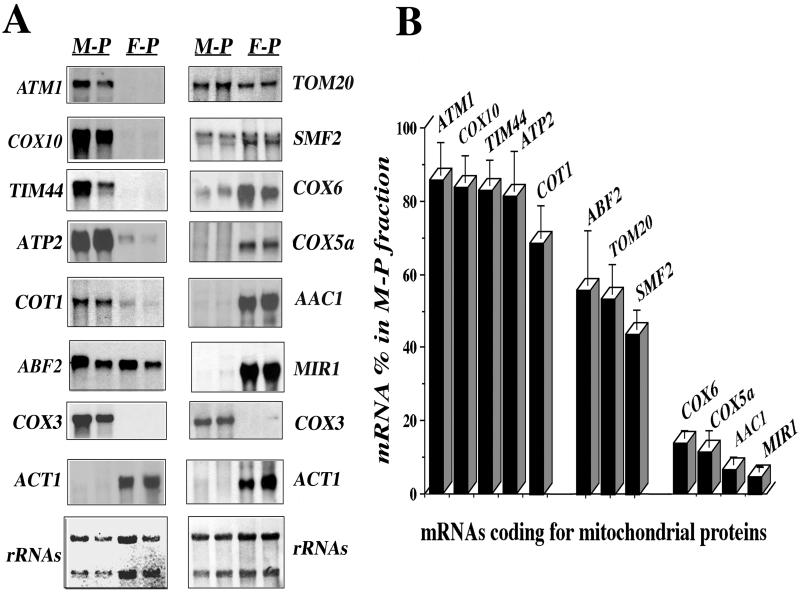
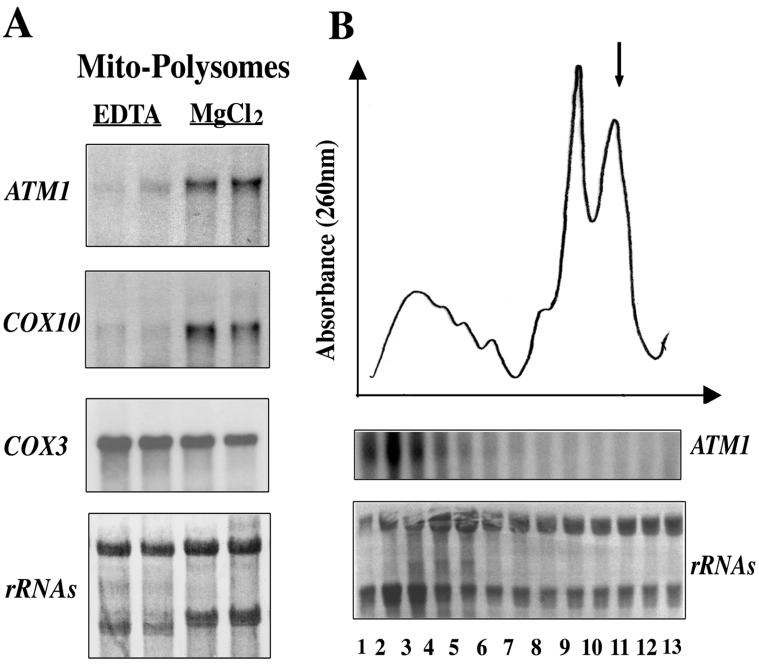
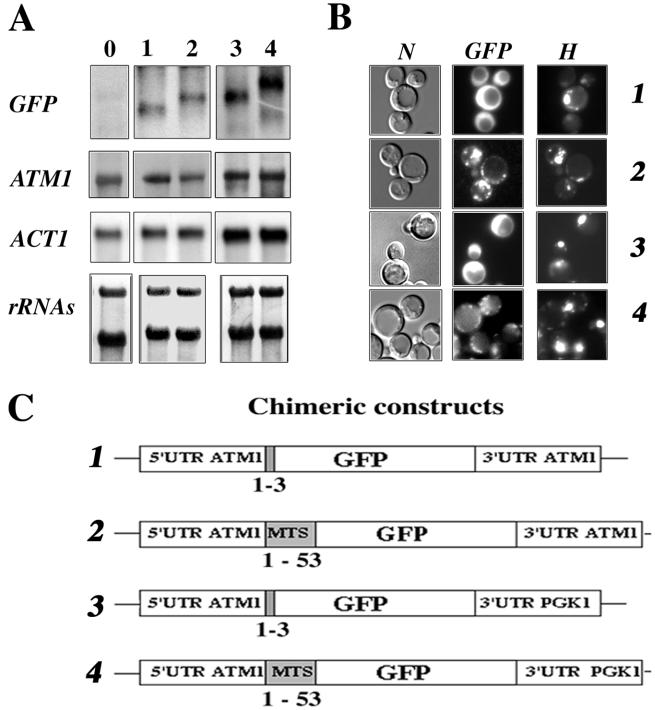
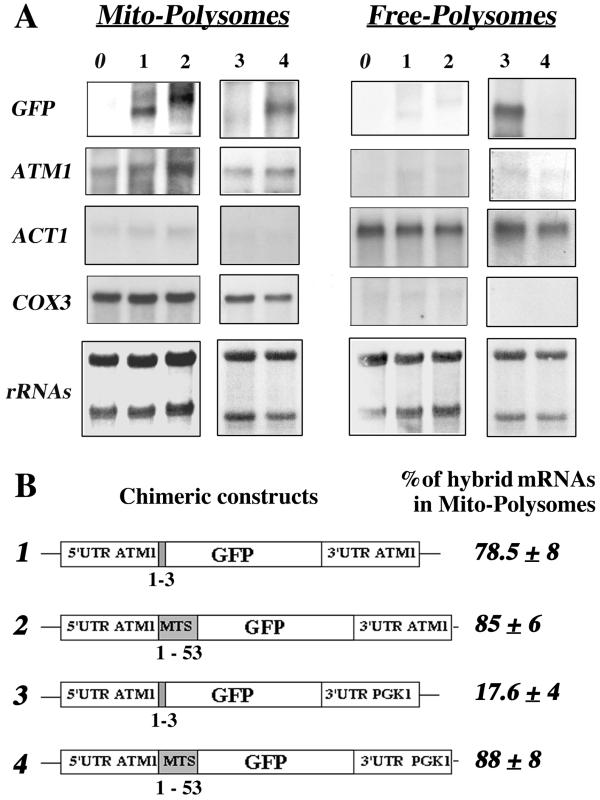
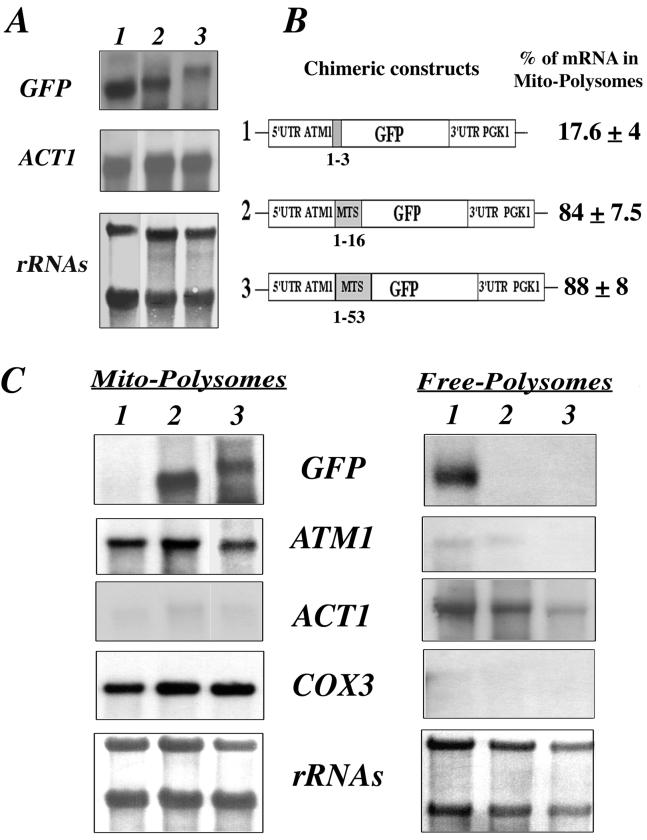
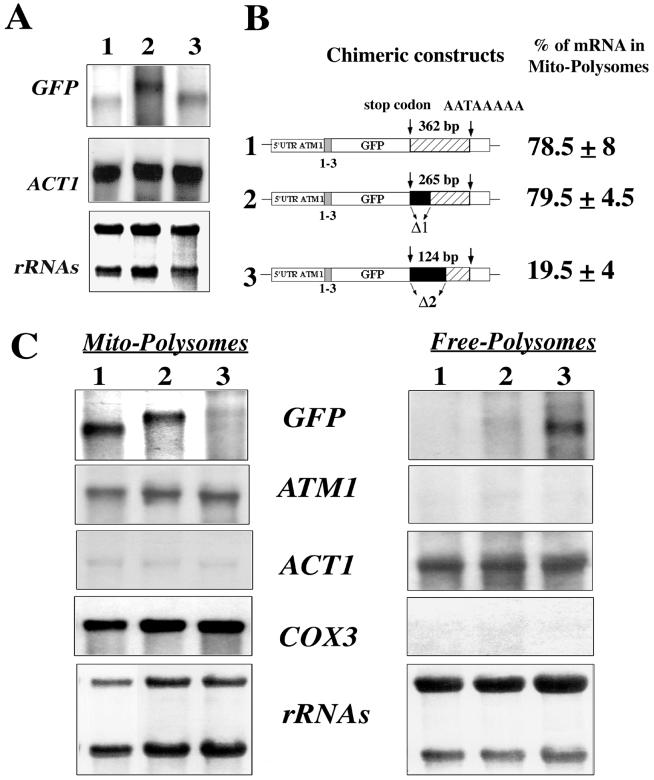
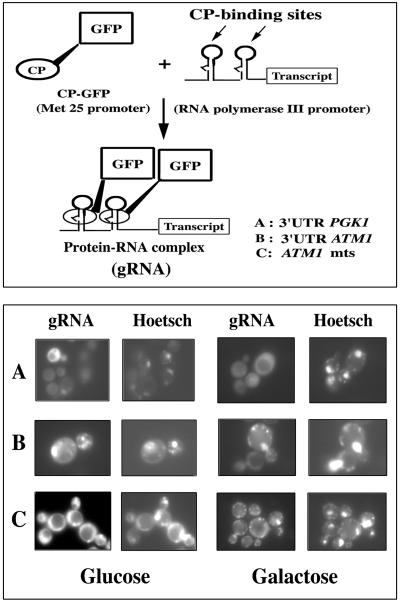
We isolated mitochondria from Saccharomyces cerevisiae to selectively study polysomes bound to the mitochondrial surface. The distribution of several mRNAs coding for mitochondrial proteins was examined in free and mitochondrion-bound polysomes. Some mRNAs exclusively localize to mitochondrion-bound polysomes, such as the ones coding for Atm1p, Cox10p, Tim44p, Atp2p, and Cot1p. In contrast, mRNAs encoding Cox6p, Cox5a, Aac1p, and Mir1p are found enriched in free cytoplasmic polysome fractions. Aac1p and Mir1p are transporters that lack cleavable presequences. Sequences required for mRNA asymmetric subcellular distribution were determined by analyzing the localization of reporter mRNAs containing the presequence coding region and/or the 3'-untranslated region (3'UTR) of ATM1, a gene encoding an ABC transporter of the mitochondrial inner membrane. Biochemical analyses of mitochondrion-bound polysomes and direct visualization of RNA localization in living yeast cells allowed us to demonstrate that either the presequence coding region or the 3'UTR of ATM1 is sufficient to allow the reporter mRNA to localize to the vicinity of the mitochondrion, independently of its translation. These data demonstrate that mRNA localization is one of the mechanisms used, in yeast, for segregating mitochondrial proteins.
Figures
Similar articles
-
Import of proteins into mitochondria. Translatable mRNAs for imported mitochondrial proteins are present in free as well as mitochondria-bound cytoplasmic polysomes.J Biol Chem. 1982 Nov 10;257(21):13048-55. J Biol Chem. 1982. PMID: 6752146
-
Identification of a human mitochondrial ABC transporter, the functional orthologue of yeast Atm1p.FEBS Lett. 1998 Dec 18;441(2):266-70. doi: 10.1016/s0014-5793(98)01560-9. FEBS Lett. 1998. PMID: 9883897
-
The ABC transporter Atm1p is required for mitochondrial iron homeostasis.FEBS Lett. 1997 Dec 1;418(3):346-50. doi: 10.1016/s0014-5793(97)01414-2. FEBS Lett. 1997. PMID: 9428742
-
Mitochondrial ABC transporters.Res Microbiol. 2001 Apr-May;152(3-4):331-40. doi: 10.1016/s0923-2508(01)01204-9. Res Microbiol. 2001. PMID: 11421280 Review.
-
Structures and functions of mitochondrial ABC transporters.Biochem Soc Trans. 2015 Oct;43(5):943-51. doi: 10.1042/BST20150118. Biochem Soc Trans. 2015. PMID: 26517908 Review.
Cited by
-
ASH1 mRNA localization in three acts.Mol Biol Cell. 2001 Sep;12(9):2567-77. doi: 10.1091/mbc.12.9.2567. Mol Biol Cell. 2001. PMID: 11553699 Free PMC article.
-
Characterization of the nuclear and cytosolic transcriptomes in human brain tissue reveals new insights into the subcellular distribution of RNA transcripts.Sci Rep. 2021 Feb 18;11(1):4076. doi: 10.1038/s41598-021-83541-1. Sci Rep. 2021. PMID: 33603054 Free PMC article.
-
Genome-wide analysis of mRNAs targeted to yeast mitochondria.EMBO Rep. 2002 Feb;3(2):159-64. doi: 10.1093/embo-reports/kvf025. Epub 2002 Jan 29. EMBO Rep. 2002. PMID: 11818335 Free PMC article.
-
Interplay between Mitochondrial Protein Import and Respiratory Complexes Assembly in Neuronal Health and Degeneration.Life (Basel). 2021 May 11;11(5):432. doi: 10.3390/life11050432. Life (Basel). 2021. PMID: 34064758 Free PMC article. Review.
-
Using the Yeast Three-Hybrid System to Identify Proteins that Interact with a Phloem-Mobile mRNA.Front Plant Sci. 2012 Aug 27;3:189. doi: 10.3389/fpls.2012.00189. eCollection 2012. Front Plant Sci. 2012. PMID: 22969782 Free PMC article.
References
-
- Achleitner G, Gaigg B, Krasser A, Kainersdorfer E, Kohlwein S D, Perktold A, Zellnig G, Daum G. Association between the endoplasmic reticulum and mitochondria of yeast facilitates interorganelle transport of phospholipids through membrane contact. Eur J Biochem. 1999;264:545–553. - PubMed
-
- Ades I Z, Butow R A. The products of mitochondria-bound cytoplasmic polysomes in yeast. J Biol Chem. 1980;255:9918–9924. - PubMed
-
- Ades I Z, Butow R A. The transport of proteins into yeast mitochondria. J Biol Chem. 1980;255:9925–9935. - PubMed
-
- Banroques J, Delahodde A, Jacq C. A mitochondrial RNA maturase gene transferred to the yeast nucleus can control mitochondrial mRNA splicing. Cell. 1986;46:837–844. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases