

Responses of sugar beet roots to iron deficiency. Changes in carbon assimilation and oxygen use
- PMID: 11027736
- PMCID: PMC59192
- DOI: 10.1104/pp.124.2.885
Responses of sugar beet roots to iron deficiency. Changes in carbon assimilation and oxygen use
Abstract
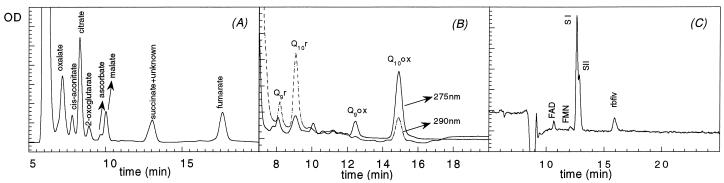
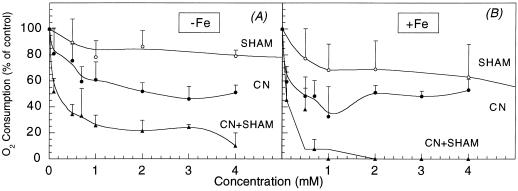
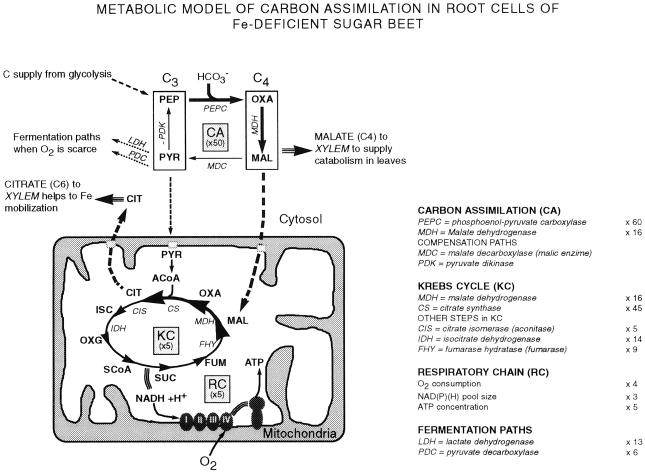
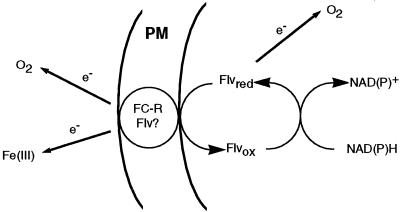
Different root parts with or without increased iron-reducing activities have been studied in iron-deficient and iron-sufficient control sugar beet (Beta vulgaris L. Monohil hybrid). The distal root parts of iron-deficient plants, 0 to 5 mm from the root apex, were capable to reduce Fe(III)-chelates and contained concentrations of flavins near 700 microM, two characteristics absent in the 5 to 10 mm sections of iron-deficient plants and the whole root of iron-sufficient plants. Flavin-containing root tips had large pools of carboxylic acids and high activities of enzymes involved in organic acid metabolism. In iron-deficient yellow root tips there was a large increase in carbon fixation associated to an increase in phosphoenolpyruvate carboxylase activity. Part of this carbon was used, through an increase in mitochondrial activity, to increase the capacity to produce reducing power, whereas another part was exported via xylem. Root respiration was increased by iron deficiency. In sugar beet iron-deficient roots flavins would provide a suitable link between the increased capacity to produce reduced nucleotides and the plasma membrane associated ferric chelate reductase enzyme(s). Iron-deficient roots had a large oxygen consumption rate in the presence of cyanide and hydroxisalycilic acid, suggesting that the ferric chelate reductase enzyme is able to reduce oxygen in the absence of Fe(III)-chelates.
Figures
References
-
- Alhendawi RA, Römheld V, Kirby EA, Marschner H. Influence of increasing bicarbonate concentrations on plant growth, organic acid accumulation in roots and iron uptake by barley, sorghum and maize. J Plant Nutr. 1997;20:1731–1753.
-
- Bedri AA, Wallace A, Rhoads WA. Assimilation of bicarbonate by roots of different plant species. Soil Sci. 1960;89:257–263.
-
- Bergmeyer HU, Gawwehn K, Grassl M. Enzymes as biochemical reagents. In: Bergmeyer HU, editor. Methods of Enzymatic Analysis. New York: Academic Press; 1974. pp. 425–556.
-
- Bialzyk J, Lechowski L. Absorption of HCO3− by roots and its effect on carbon metabolism of tomato. J Plant Nutr. 1992;15:293–312.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
