

Characterization of the hydrogen-deuterium exchange activities of the energy-transducing HupSL hydrogenase and H(2)-signaling HupUV hydrogenase in Rhodobacter capsulatus
- PMID: 11029418
- PMCID: PMC94732
- DOI: 10.1128/JB.182.21.5997-6004.2000
Characterization of the hydrogen-deuterium exchange activities of the energy-transducing HupSL hydrogenase and H(2)-signaling HupUV hydrogenase in Rhodobacter capsulatus
Abstract
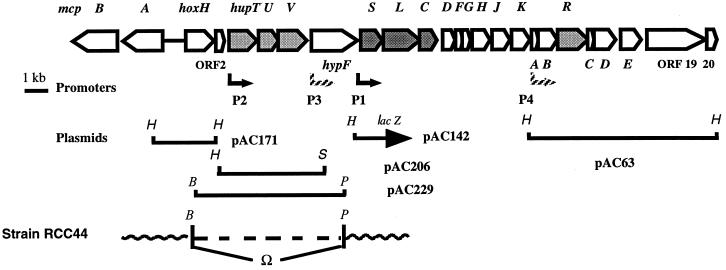
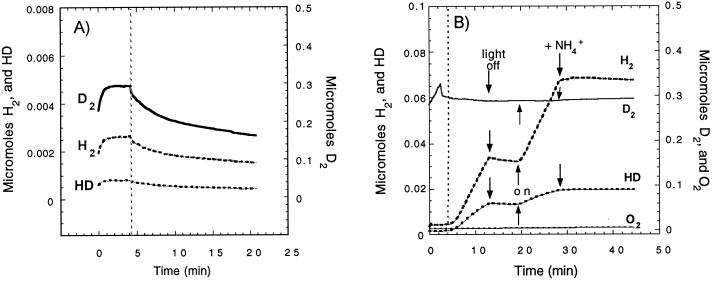
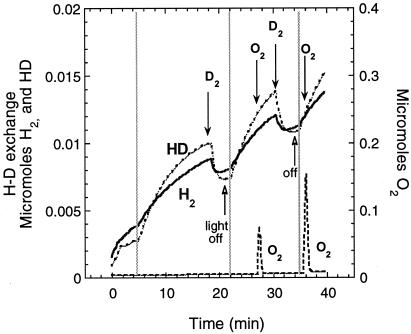
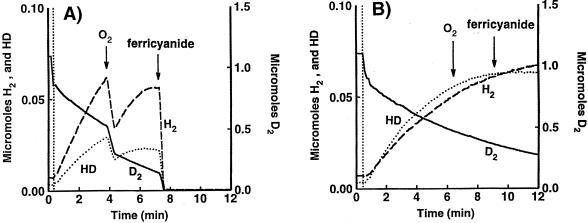
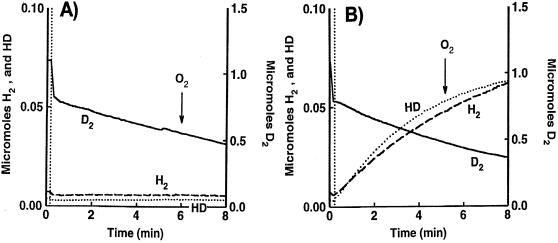
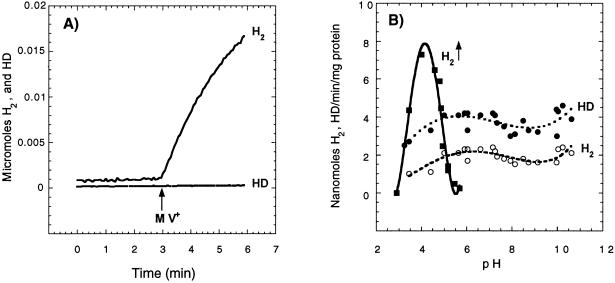
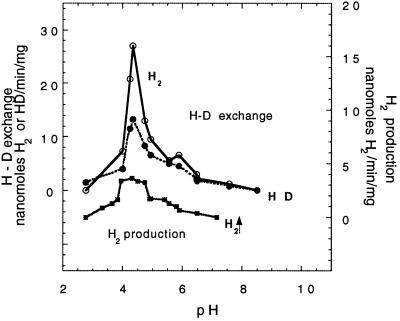
Rhodobacter capsulatus synthesizes two homologous protein complexes capable of activating molecular H(2), a membrane-bound [NiFe] hydrogenase (HupSL) linked to the respiratory chain, and an H(2) sensor encoded by the hupUV genes. The activities of hydrogen-deuterium (H-D) exchange catalyzed by the hupSL-encoded and the hupUV-encoded enzymes in the presence of D(2) and H(2)O were studied comparatively. Whereas HupSL is in the membranes, HupUV activity was localized in the soluble cytoplasmic fraction. Since the hydrogenase gene cluster of R. capsulatus contains a gene homologous to hoxH, which encodes the large subunit of NAD-linked tetrameric soluble hydrogenases, the chromosomal hoxH gene was inactivated and hoxH mutants were used to demonstrate the H-D exchange activity of the cytoplasmic HupUV protein complex. The H-D exchange reaction catalyzed by HupSL hydrogenase was maximal at pH 4. 5 and inhibited by acetylene and oxygen, whereas the H-D exchange catalyzed by the HupUV protein complex was insensitive to acetylene and oxygen and did not vary significantly between pH 4 and pH 11. Based on these properties, the product of the accessory hypD gene was shown to be necessary for the synthesis of active HupUV enzyme. The kinetics of HD and H(2) formed in exchange with D(2) by HupUV point to a restricted access of protons and gasses to the active site. Measurement of concentration changes in D(2), HD, and H(2) by mass spectrometry showed that, besides the H-D exchange reaction, HupUV oxidized H(2) with benzyl viologen, produced H(2) with reduced methyl viologen, and demonstrated true hydrogenase activity. Therefore, not only with respect to its H(2) signaling function in the cell, but also to its catalytic properties, the HupUV enzyme represents a distinct class of hydrogenases.
Figures
References
-
- Appel J, Schulz R. Hydrogen metabolism in organisms with oxygenic photosynthesis: hydrogenases as important regulatory devices for a proper redox poising? J Photochem Photobiol. 1998;47:1–11.
-
- Cauvin B, Colbeau A, Vignais P M. The hydrogenase structural operon in Rhodobacter capsulatus contains a third gene, hupM, necessary for the formation of a physiologically competent hydrogenase. Mol Microbiol. 1991;5:2519–2527. - PubMed
-
- Colbeau A, Elsen S, Tomiyama M, Zorin N A, Dimon B, Vignais P M. Rhodobacter capsulatus HypF is involved in regulation of hydrogenase synthesis through the HupUV proteins. Eur J Biochem. 1998;251:65–71. - PubMed
-
- Colbeau A, Godfroy A, Vignais P M. Cloning of DNA fragments carrying hydrogenase genes of Rhodopseudomonas capsulata. Biochimie. 1986;68:147–155. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
