

Defects in D-alanyl-lipoteichoic acid synthesis in Streptococcus mutans results in acid sensitivity
- PMID: 11029425
- PMCID: PMC94739
- DOI: 10.1128/JB.182.21.6055-6065.2000
Defects in D-alanyl-lipoteichoic acid synthesis in Streptococcus mutans results in acid sensitivity
Abstract
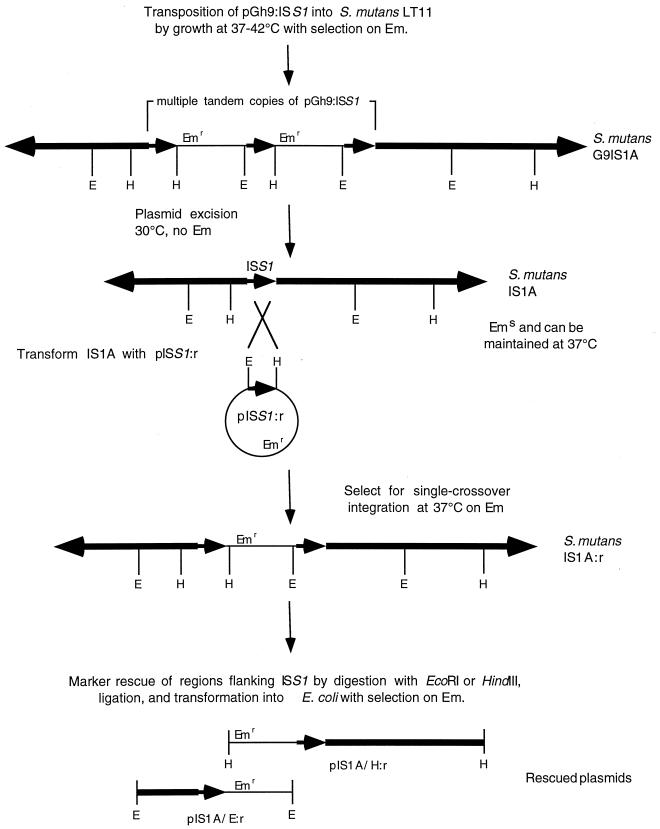
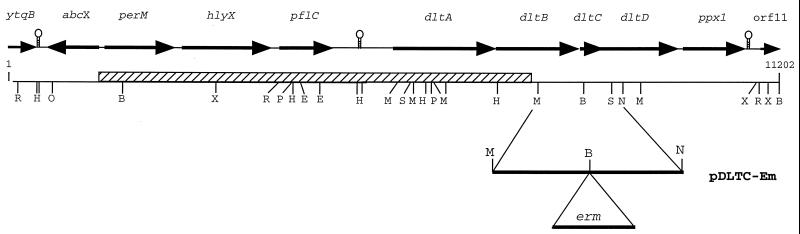
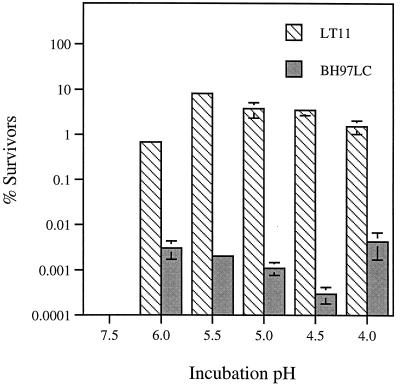
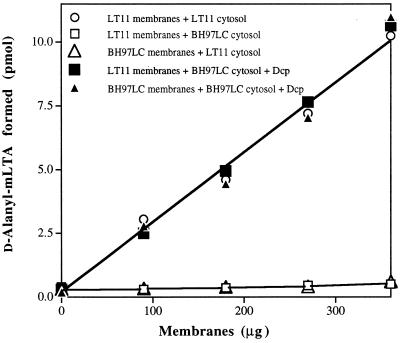
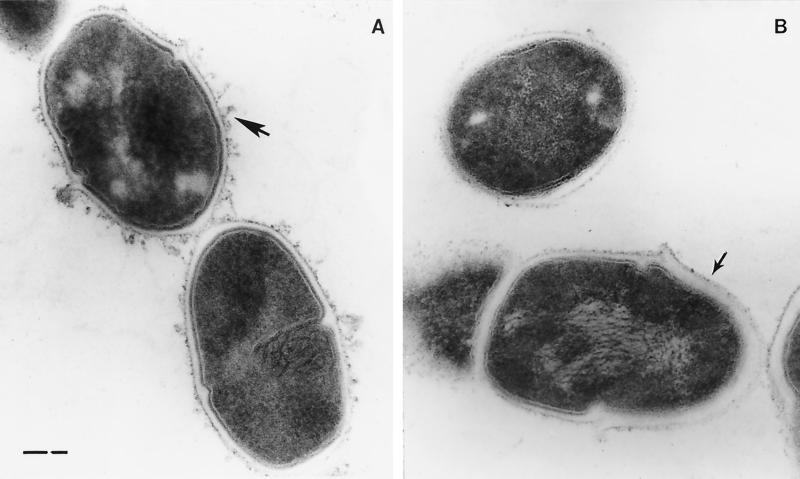
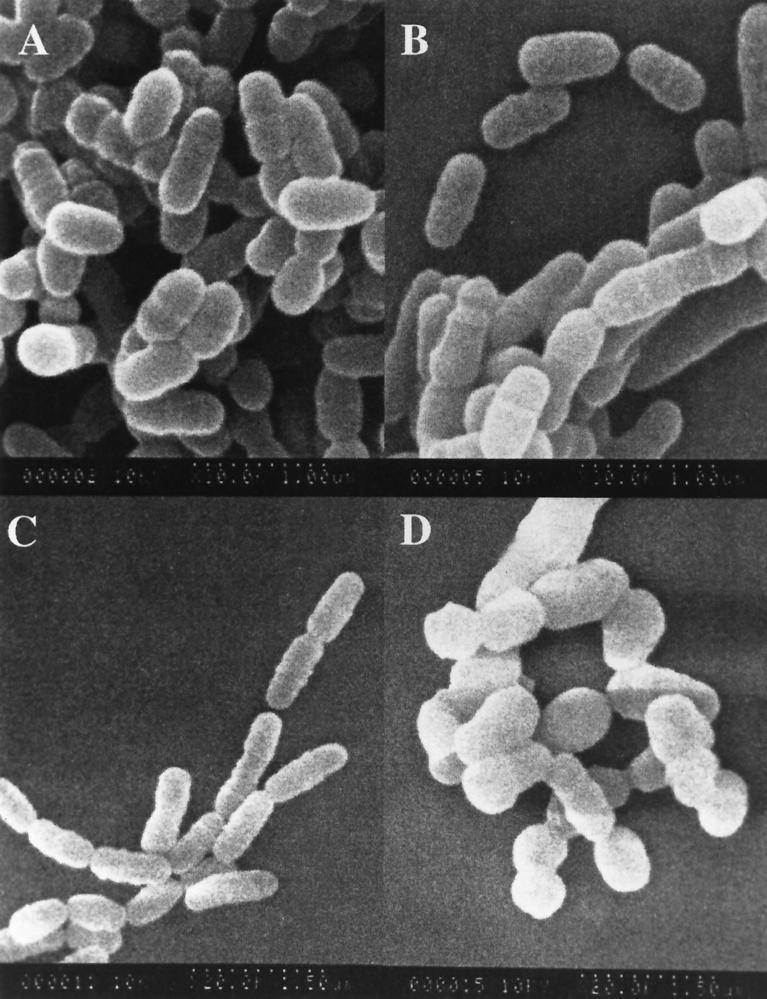
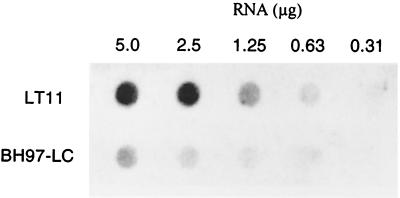
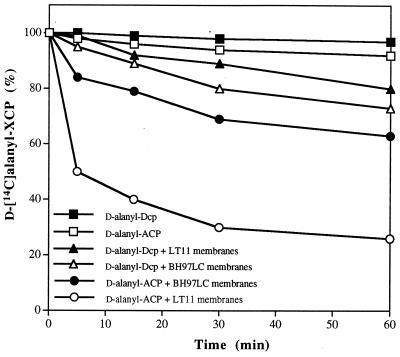
In the cariogenic organism, Streptococcus mutans, low pH induces an acid tolerance response (ATR). To identify acid-regulated proteins comprising the ATR, transposon mutagenesis with the thermosensitive plasmid pGh9:ISS1 was used to produce clones that were able to grow at neutral pH, but not in medium at pH 5.0. Sequence analysis of one mutant (IS1A) indicated that transposition had created a 6.3-kb deletion, one end of which was in dltB of the dlt operon encoding four proteins (DltA-DltD) involved in the synthesis of D-alanyl-lipoteichoic acid. Inactivation of the dltC gene, encoding the D-alanyl carrier protein (Dcp), resulted in the generation of the acid-sensitive mutant, BH97LC. Compared to the wild-type strain, LT11, the mutant exhibited a threefold-longer doubling time and a 33% lower growth yield. In addition, it was unable to initiate growth below pH 6.5 and unadapted cells were unable to survive a 3-h exposure in medium buffered at pH 3.5, while a pH of 3.0 was required to kill the wild type in the same time period. Also, induction of the ATR in BH97LC, as measured by the number of survivors at a pH killing unadapted cells, was 3 to 4 orders of magnitude lower than that exhibited by the wild type. While the LTA of both strains contained a similar average number of glycerolphosphate residues, permeabilized cells of BH97LC did not incorporate D-[(14)C]alanine into this amphiphile. This defect was correlated with the deficiency of Dcp. Chemical analysis of the LTA purified from the mutant confirmed the absence of D-alanine-esters. Electron micrographs showed that BH97LC is characterized by unequal polar caps and is devoid of a fibrous extracellular matrix present on the surface of the wild-type cells. Proton permeability assays revealed that the mutant was more permeable to protons than the wild type. This observation suggests a mechanism for the loss of the characteristic acid tolerance response in S. mutans.
Figures
References
-
- Archibald A R, Baddiley J, Heptinstall S. The alanine ester content and magnesium binding capacity of walls of Staphylococcus aureus H grown at different pH values. Biochim Biophys Acta. 1973;291:629–634. - PubMed
-
- Bleiweis A S, Craig R C. A model system for the purification of amphophiles: the preparation of lipoteichoic acid using liposomes. In: Shockman G D, Wicken A J, editors. Chemistry and biological activities of bacterial surface amphophiles. New York, N.Y: Academic Press; 1981. pp. 43–49.
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
