

Direct association of ligand-binding and pore domains in homo- and heterotetrameric inositol 1,4,5-trisphosphate receptors
- PMID: 11032812
- PMCID: PMC313997
- DOI: 10.1093/emboj/19.20.5450
Direct association of ligand-binding and pore domains in homo- and heterotetrameric inositol 1,4,5-trisphosphate receptors
Abstract
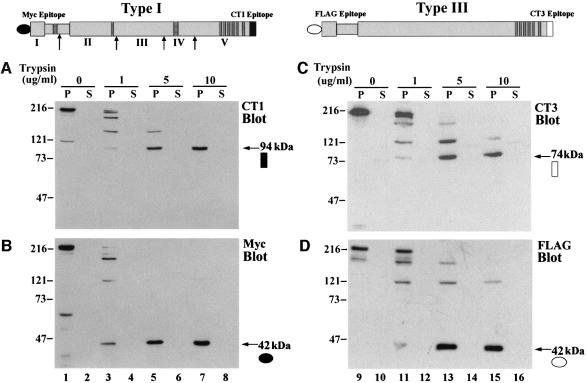
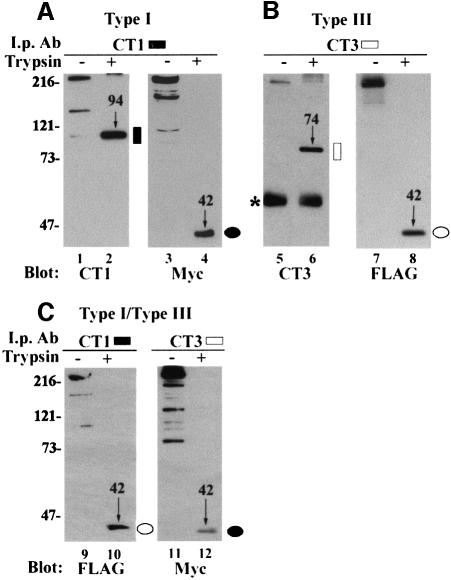
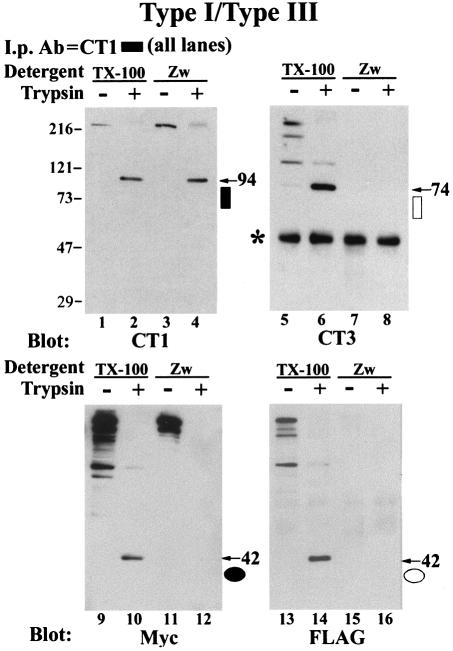
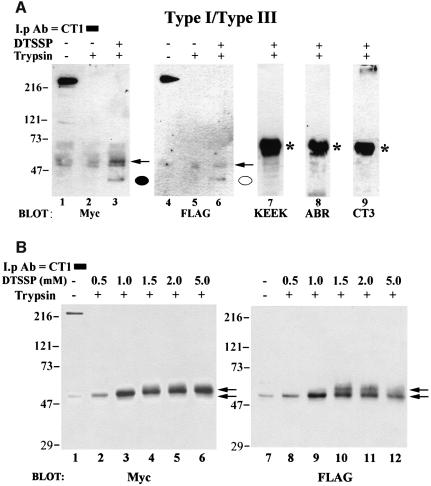
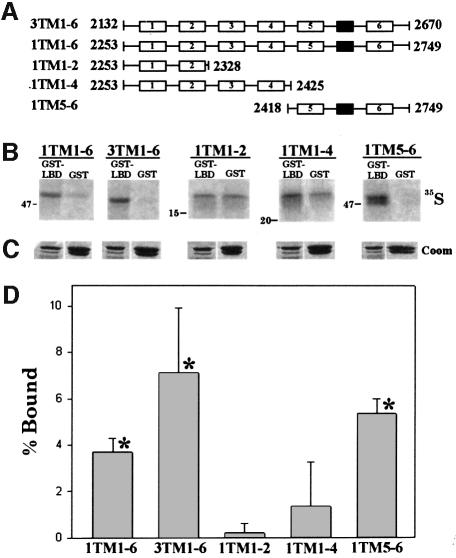
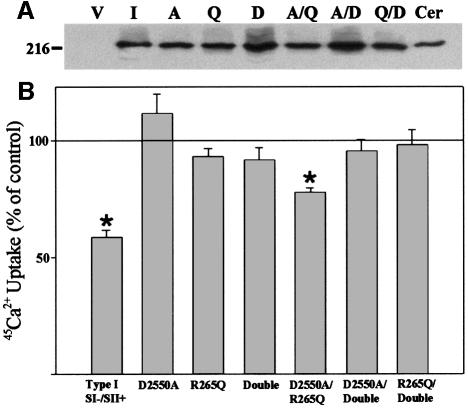
Inositol 1,4,5-trisphosphate receptors (IP(3)Rs) are a family of intracellular Ca(2+) channels that exist as homo- or heterotetramers. In order to determine whether the N-terminal ligand-binding domain is in close physical proximity to the C-terminal pore domain, we prepared microsomal membranes from COS-7 cells expressing recombinant type I and type III IP(3)R isoforms. Trypsin digestion followed by cross-linking and co-immunoprecipitation of peptide fragments suggested an inter-subunit N- and C-terminal interaction in both homo- and heterotetramers. This observation was further supported by the ability of in vitro translated C-terminal peptides to interact specifically with an N-terminal fusion protein. Using a (45)Ca(2+) flux assay, we provide functional evidence that the ligand-binding domain of one subunit can gate the pore domain of an adjacent subunit. We conclude that common structural motifs are shared between the type I and type III IP(3)Rs and propose that the gating mechanism of IP(3)R Ca(2+) channels involves the association of the N-terminus of one subunit with the C-terminus of an adjacent subunit in both homo- and heterotetrameric complexes.
Figures
References
-
- Berridge M.J. and Irvine,R.F. (1989) Inositol phosphates and cell signaling. Nature, 341, 197–205. - PubMed
-
- Blondel O., Takeda,J., Janssen,H., Seino,S. and Bell,G. (1993) Sequence and functional characterization of a third inositol trisphosphate receptor subtype, IP3R-3, expressed in pancreatic islets, kidney, gastrointestinal tract and other tissues. J. Biol. Chem., 268, 11356–11363. - PubMed
-
- Boehning D. and Joseph,S.K. (2000) Functional properties of recombinant type I and type III inositol 1,4,5-trisphosphate receptor isoforms expressed in COS-7 cells. J. Biol. Chem., 275, 21492–21499. - PubMed
-
- Bronfman M., Loyola,G. and Koenig,C.S. (1998) Isolation of intact organelles by differential centrifugation of digitonin-treated hepatocytes using a table Eppendorf centrifuge. Anal. Biochem., 255, 252–256. - PubMed
-
- Dufour J., Arias,I.M. and Turner,T.J. (1997) Inositol 1,4,5-trisphosphate and calcium regulate the calcium channel function of the hepatic inositol 1,4,5-trisphosphate receptor. J. Biol. Chem., 272, 2675–2681. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
