

Neisseria gonorrhoeae porin modifies the oxidative burst of human professional phagocytes
- PMID: 11035728
- PMCID: PMC97702
- DOI: 10.1128/IAI.68.11.6215-6222.2000
Neisseria gonorrhoeae porin modifies the oxidative burst of human professional phagocytes
Abstract
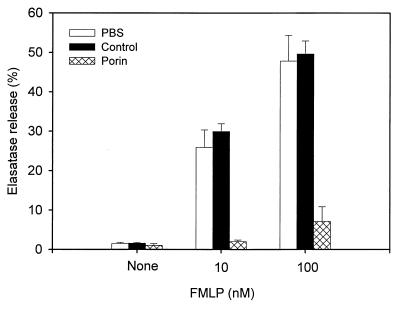
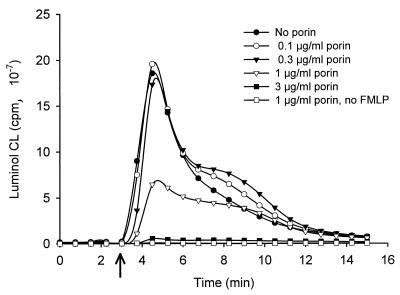
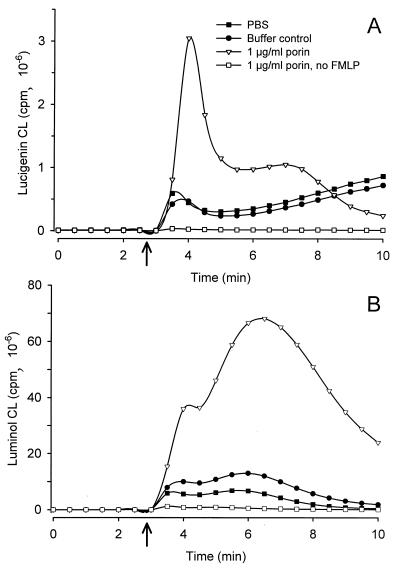
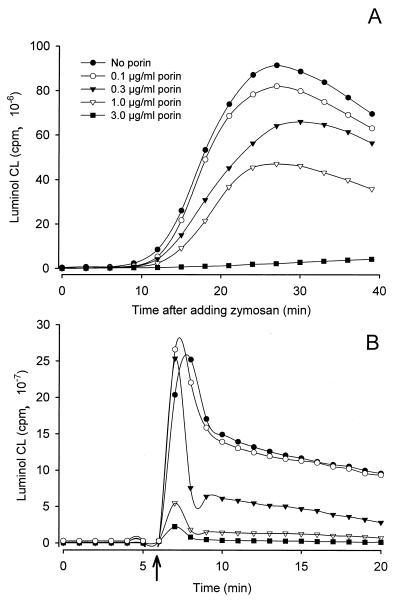
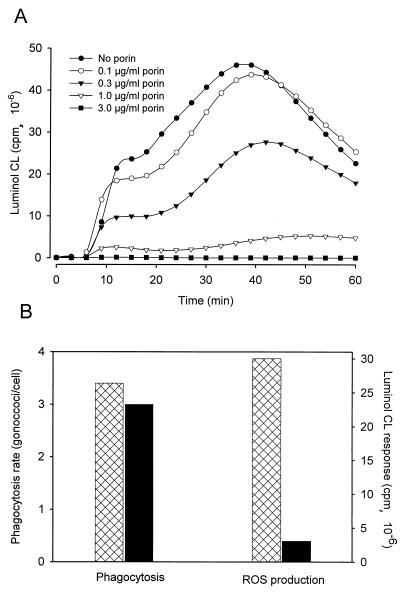
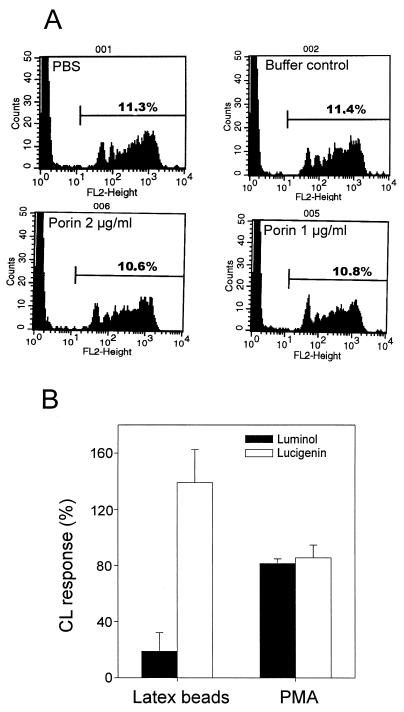
A hallmark of infection with the gram-negative bacterium Neisseria gonorrhoeae is the local infiltration and subsequent activation of polymorphonuclear neutrophils. Several gonococcal outer membrane proteins are involved in the interaction with and the activation of these phagocytes, including gonococcal porin, the most abundant protein in the outer membrane. Previous work suggests that this porin plays a role in various cellular processes, including inhibiting neutrophils activation and phagosome maturation in professional phagocytes. Here we investigated the ability of porin to modify the oxidative metabolism of human peripheral blood neutrophils and monocytes in response to particulate stimuli (including live gonococci) and soluble agents. The activation of the oxidative metabolism was determined by chemiluminescence amplified with either luminol or lucigenin. We found that treatment of the phagocytes with porin inhibits the release of reactive oxygen species measured as luminol-enhanced chemiluminescence in response to zymosan, latex particles, and gonococci. The engulfment of these particles was not, however, affected by porin treatment. Similar effects of porin on the chemiluminescence response were observed in cytochalasin B-treated neutrophils exposed to the soluble chemotactic peptide N-formylmethionyl-leucyl-phenylalanine. This indicates that porin selectively inhibits granule fusion with those cellular membranes that are in direct contact with porin, namely, the phagosomal and plasma membranes. This porin-induced downregulation of oxidative metabolism may be a potent mechanism by which gonococci modulate oxygen-dependent reactions by activated phagocytes at inflammation sites.
Figures
References
-
- Aida Y, Pabst M J. Priming of neutrophils by lipopolysaccharide for enhanced release of superoxide. Requirement for plasma but not for tumor necrosis factor-alpha. J Immunol. 1990;145:3017–3030. - PubMed
-
- Albrecht D, Jungi T W. Luminol-enhanced chemiluminescence induced in peripheral blood-derived human phagocytes: obligatory requirement of myeloperoxidase exoocytosis by monocytes. J Leukoc Biol. 1993;54:300–306. - PubMed
-
- Allen R C. Phagocytic leukocyte activities and chemiluminescence: a kinetic approach to analysis. Methods Enzymol. 1986;133:449–493. - PubMed
-
- Bauer F J, Rudel T, Stein M, Meyer T F. Mutagenesis of the Neisseria gonorrhoeae porin reduces invasion in epithelial cells and enhances phagocyte responsiveness. Mol Microbiol. 1999;31:903–913. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
