

Frataxin activates mitochondrial energy conversion and oxidative phosphorylation
- PMID: 11035806
- PMCID: PMC17325
- DOI: 10.1073/pnas.220403797
Frataxin activates mitochondrial energy conversion and oxidative phosphorylation
Abstract
Friedreich's ataxia (FA) is an autosomal recessive disease caused by decreased expression of the mitochondrial protein frataxin. The biological function of frataxin is unclear. The homologue of frataxin in yeast, YFH1, is required for cellular respiration and was suggested to regulate mitochondrial iron homeostasis. Patients suffering from FA exhibit decreased ATP production in skeletal muscle. We now demonstrate that overexpression of frataxin in mammalian cells causes a Ca(2+)-induced up-regulation of tricarboxylic acid cycle flux and respiration, which, in turn, leads to an increased mitochondrial membrane potential (delta psi(m)) and results in an elevated cellular ATP content. Thus, frataxin appears to be a key activator of mitochondrial energy conversion and oxidative phosphorylation.
Figures
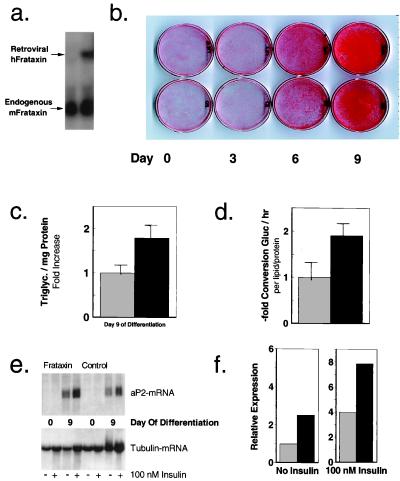
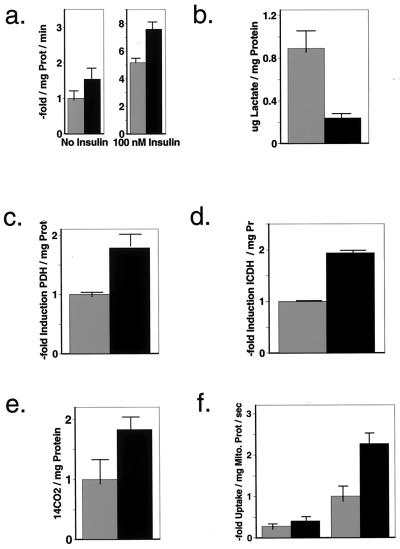
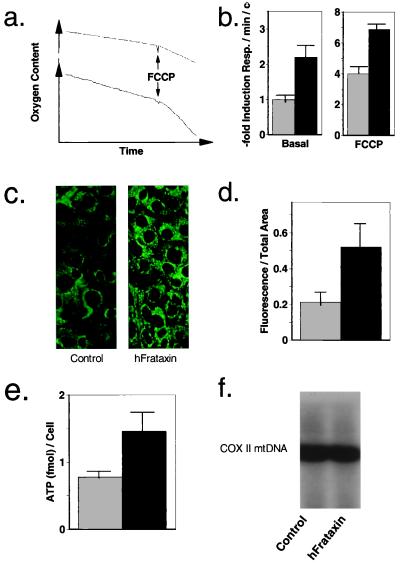
References
-
- Friedreich N. Virchows Arch Pathol Anat Physiol Klin Med. 1863;26:391–419.
-
- Finocchiaro G, Baio G, Micossi P, Pozza G, di Donato S. Neurology. 1988;38:1292–1296. - PubMed
-
- Tolis G, Mehta A, Andermann E, Harvey C, Barbeau A. Can J Neurol Sci. 1980;7:397–400. - PubMed
-
- Khan R J, Andermann E, Fantus I G. Metabolism. 1986;35:1017–1023. - PubMed
-
- Hebinck J, Hardt C, Schoels L, Vorgerd M, Briedigkeit L, Kahn C R, Ristow M. Diabetes. 2000;49:1604–1607. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
