

Dnm1p GTPase-mediated mitochondrial fission is a multi-step process requiring the novel integral membrane component Fis1p
- PMID: 11038183
- PMCID: PMC2192649
- DOI: 10.1083/jcb.151.2.367
Dnm1p GTPase-mediated mitochondrial fission is a multi-step process requiring the novel integral membrane component Fis1p
Abstract
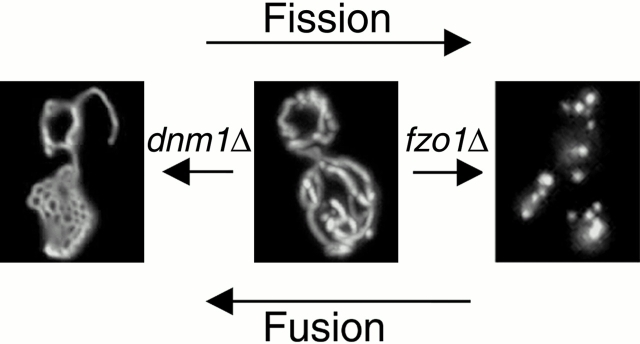
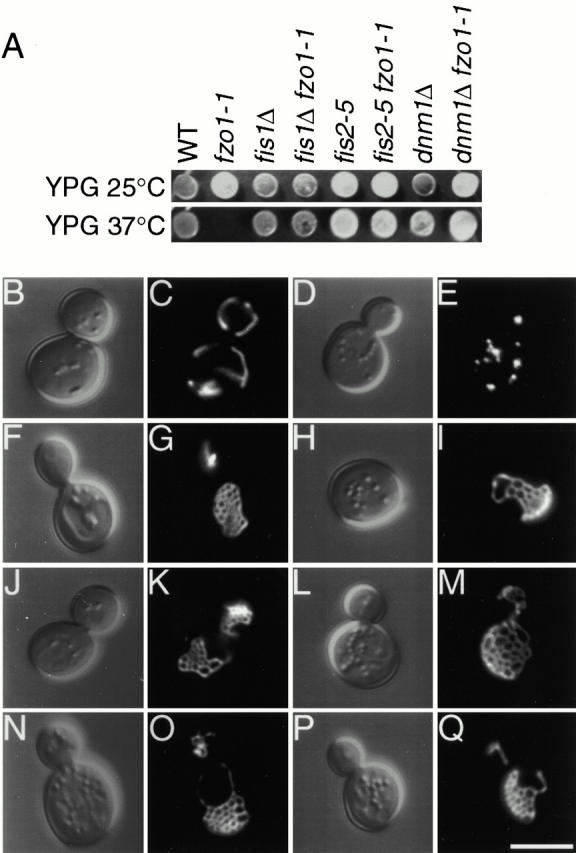
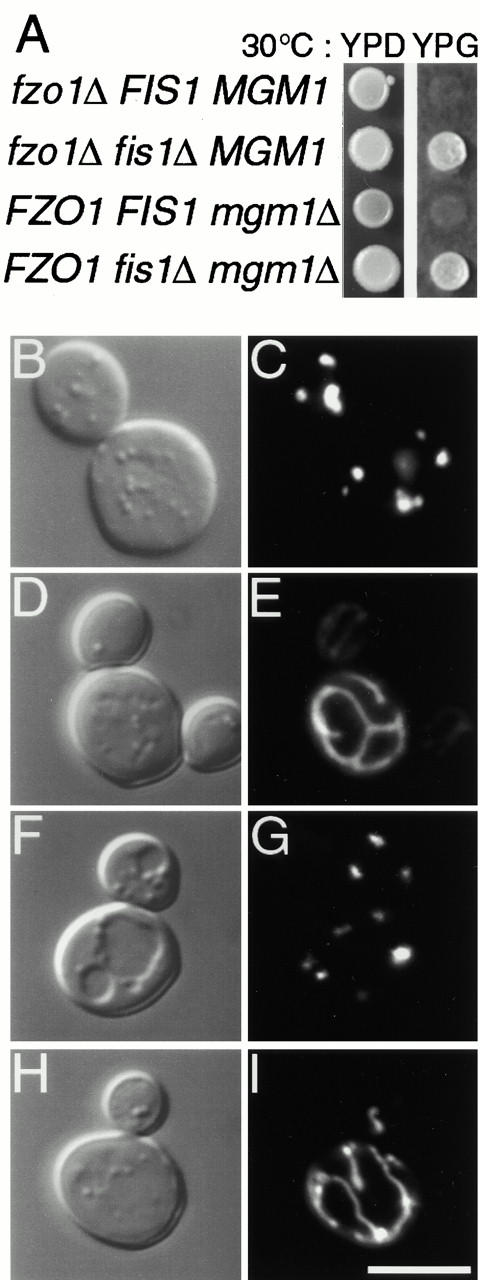
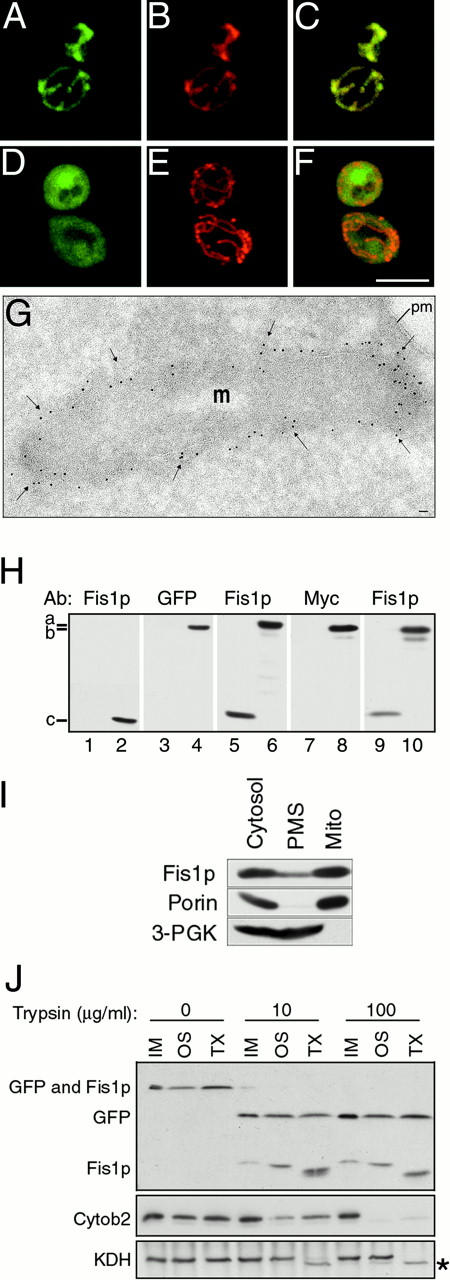
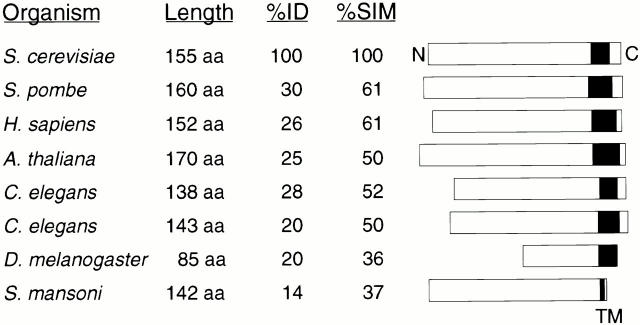
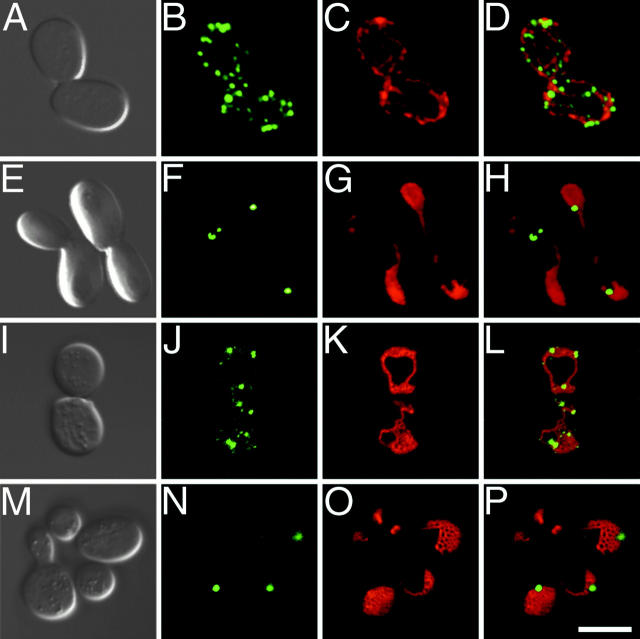
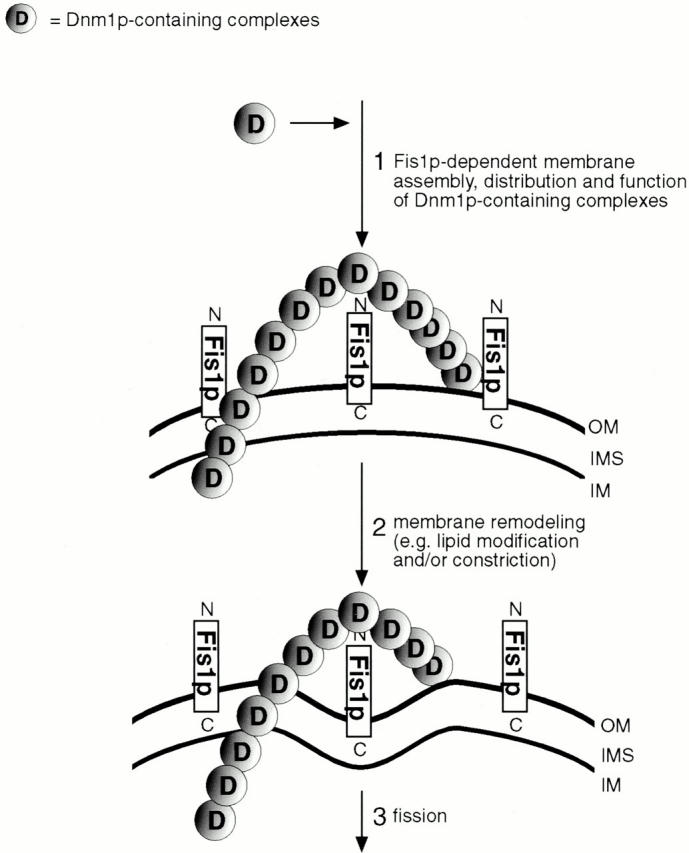
Yeast Dnm1p is a soluble, dynamin-related GTPase that assembles on the outer mitochondrial membrane at sites where organelle division occurs. Although these Dnm1p-containing complexes are thought to trigger constriction and fission, little is known about their composition and assembly, and molecules required for their membrane recruitment have not been isolated. Using a genetic approach, we identified two new genes in the fission pathway, FIS1 and FIS2. FIS1 encodes a novel, outer mitochondrial membrane protein with its amino terminus exposed to the cytoplasm. Fis1p is the first integral membrane protein shown to participate in a eukaryotic membrane fission event. In a related study (Tieu, Q., and J. Nunnari. 2000. J. Cell Biol. 151:353-365), it was shown that the FIS2 gene product (called Mdv1p) colocalizes with Dnm1p on mitochondria. Genetic and morphological evidence indicate that Fis1p, but not Mdv1p, function is required for the proper assembly and distribution of Dnm1p-containing fission complexes on mitochondrial tubules. We propose that mitochondrial fission in yeast is a multi-step process, and that membrane-bound Fis1p is required for the proper assembly, membrane distribution, and function of Dnm1p-containing complexes during fission.
Figures
Comment in
-
A mitochondrial division apparatus takes shape.J Cell Biol. 2000 Oct 16;151(2):F1-4. doi: 10.1083/jcb.151.2.f1. J Cell Biol. 2000. PMID: 11038192 Free PMC article. No abstract available.
References
-
- Adari H., Lowy D.R., Willumsen B.M., Der C.J., McCormick F. Guanosine triphosphatase activating protein (GAP) interacts with the p21 ras effector binding domain. Science. 1988;240:518–521. - PubMed
-
- Beech P.L., Gilson P.R. FtsZ and organelle division in Protists. Protist. 2000;151:11–16. - PubMed
-
- Beech P.L., Nheu T., Schultz T., Herbert S., Lithgow T., Gilson P.R., McFadden G.I. Mitochondrial FtsZ in a chromophyte alga. Science. 2000;287:1276–1279. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
