

The transmembrane domain of influenza hemagglutinin exhibits a stringent length requirement to support the hemifusion to fusion transition
- PMID: 11038188
- PMCID: PMC2192652
- DOI: 10.1083/jcb.151.2.425
The transmembrane domain of influenza hemagglutinin exhibits a stringent length requirement to support the hemifusion to fusion transition
Abstract
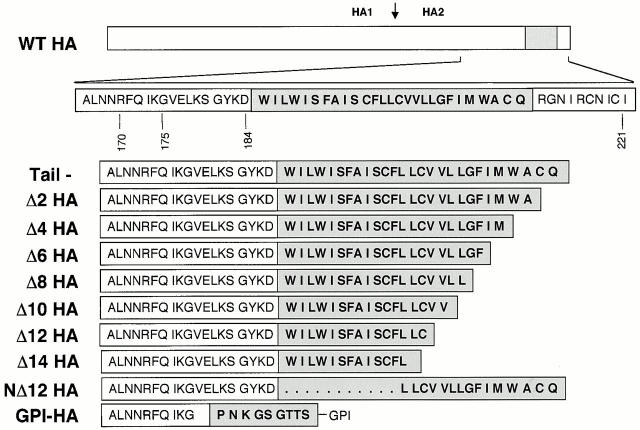
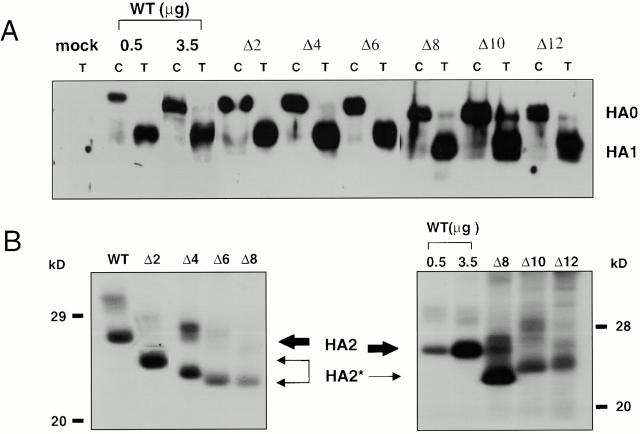
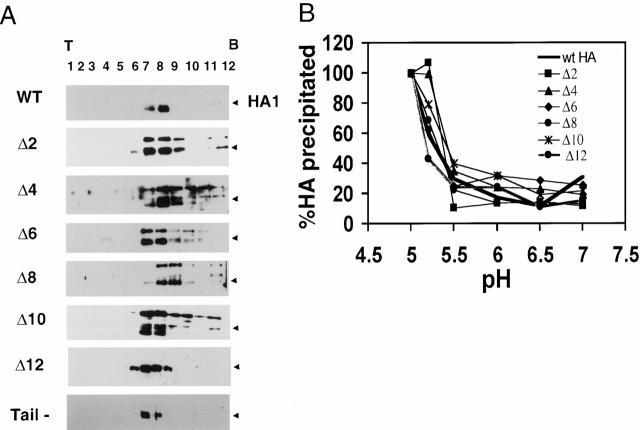
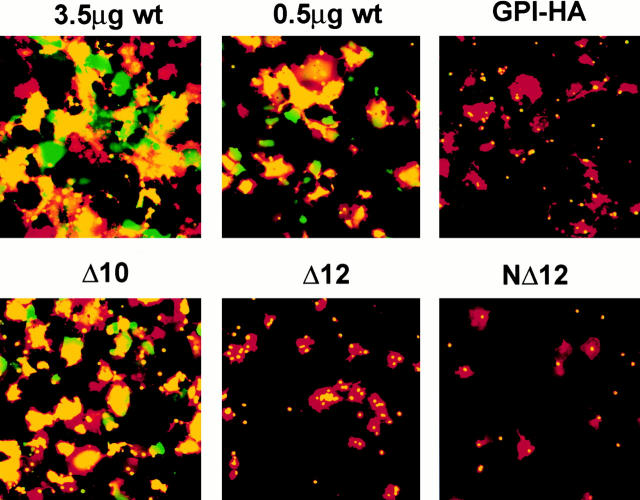
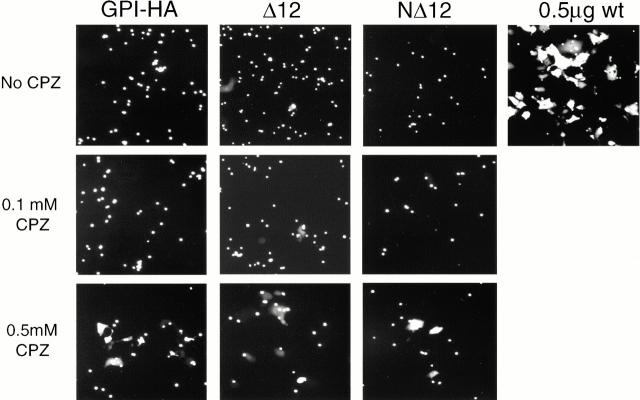
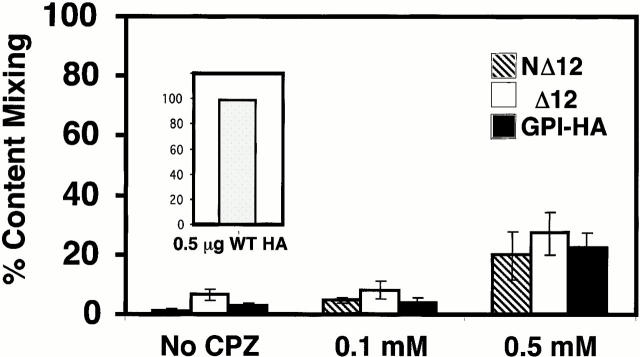
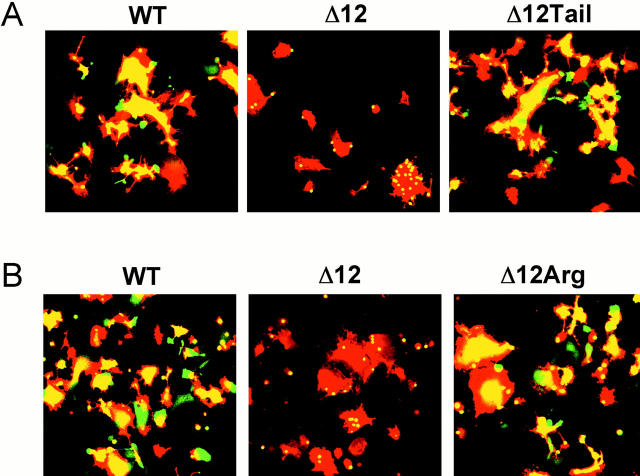
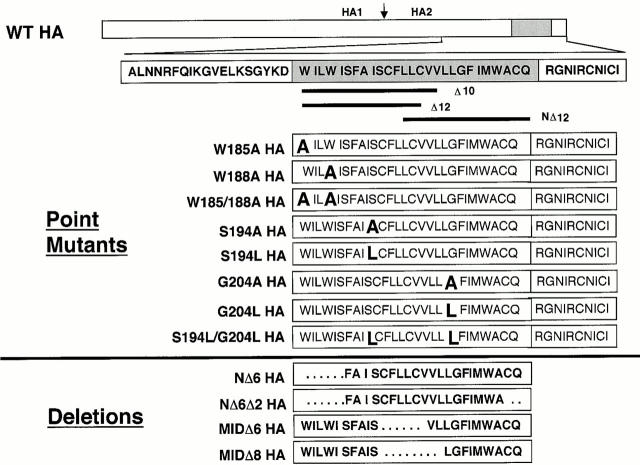
Glycosylphosphatidylinositol-anchored influenza hemagglutinin (GPI-HA) mediates hemifusion, whereas chimeras with foreign transmembrane (TM) domains mediate full fusion. A possible explanation for these observations is that the TM domain must be a critical length in order for HA to promote full fusion. To test this hypothesis, we analyzed biochemical properties and fusion phenotypes of HA with alterations in its 27-amino acid TM domain. Our mutants included sequential 2-amino acid (Delta2-Delta14) and an 11-amino acid deletion from the COOH-terminal end, deletions of 6 or 8 amino acids from the NH(2)-terminal and middle regions, and a deletion of 12 amino acids from the NH(2)-terminal end of the TM domain. We also made several point mutations in the TM domain. All of the mutants except Delta14 were expressed at the cell surface and displayed biochemical properties virtually identical to wild-type HA. All the mutants that were expressed at the cell surface promoted full fusion, with the notable exception of deletions of >10 amino acids. A mutant in which 11 amino acids were deleted was severely impaired in promoting full fusion. Mutants in which 12 amino acids were deleted (from either end) mediated only hemifusion. Hence, a TM domain of 17 amino acids is needed to efficiently promote full fusion. Addition of either the hydrophilic HA cytoplasmic tail sequence or a single arginine to Delta12 HA, the hemifusion mutant that terminates with 15 (hydrophobic) amino acids of the HA TM domain, restored full fusion activity. Our data support a model in which the TM domain must span the bilayer to promote full fusion.
Figures



Comment in
-
HIV-1 membrane fusion: targets of opportunity.J Cell Biol. 2000 Oct 16;151(2):F9-14. doi: 10.1083/jcb.151.2.f9. J Cell Biol. 2000. PMID: 11038194 Free PMC article. No abstract available.
References
-
- Bullough P.A., Hughson F.M., Skehel J.J., Wiley D.C. Structure of influenza haemagglutinin at the pH of membrane fusion. Nature. 1994;371:37–43. - PubMed
-
- Carr C.M., Kim P.S. A spring-loaded mechanism for the conformational change of influenza hemagglutinin. Cell. 1993;73:823–832. - PubMed
-
- Chernomordik L.V., Melikyan G.B., Chizmadzhev Y.A. Biomembrane fusiona new concept derived from model studies using two interacting planar lipid bilayers. Biochim. Biophys. Acta. 1987;906:309–352. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
