

The pea light-independent photomorphogenesis1 mutant results from partial duplication of COP1 generating an internal promoter and producing two distinct transcripts
- PMID: 11041887
- PMCID: PMC149130
- DOI: 10.1105/tpc.12.10.1927
The pea light-independent photomorphogenesis1 mutant results from partial duplication of COP1 generating an internal promoter and producing two distinct transcripts
Abstract
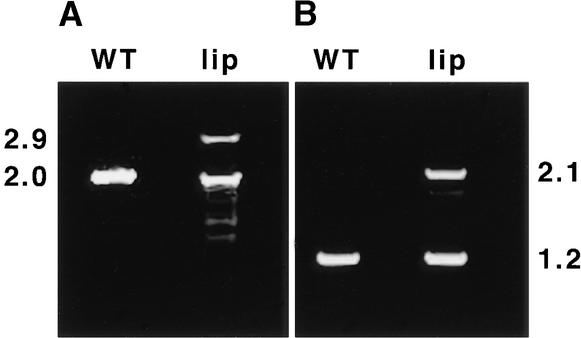
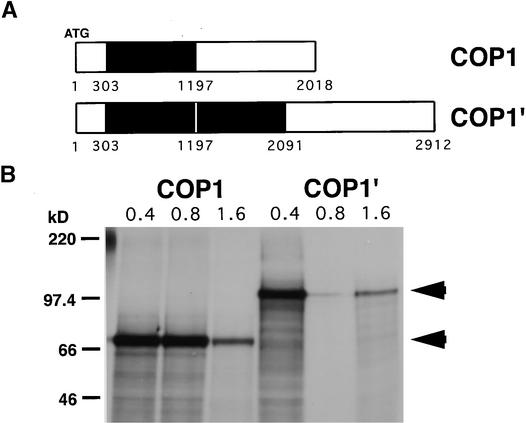
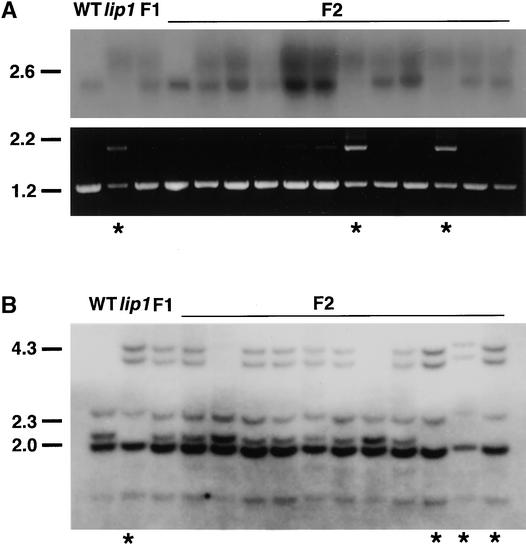
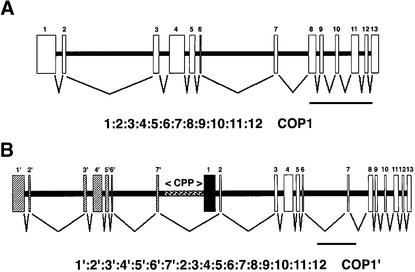
The pea lip1 (light-independent photomorphogenesis1) mutant shows many of the characteristics of light-grown development when grown in continuous darkness. To investigate the identity of LIP1, cDNAs encoding the pea homolog of COP1, a repressor of photomorphogenesis identified in Arabidopsis, were isolated from wild-type and lip1 pea seedlings. lip1 seedlings contained a wild-type COP1 transcript as well as a larger COP1' transcript that contained an internal in-frame duplication of 894 bp. The COP1' transcript segregated with the lip1 phenotype in F(2) seedlings and could be translated in vitro to produce a protein of approximately 100 kD. The COP1 gene in lip1 peas contained a 7.5-kb duplication, consisting of exons 1 to 7 of the wild-type sequence, located 2.5 kb upstream of a region of genomic DNA identical to the wild-type COP1 DNA sequence. Transcription and splicing of the mutant COP1 gene was predicted to produce the COP1' transcript, whereas transcription from an internal promoter in the 2.5-kb region of DNA located between the duplicated regions of COP1 would produce the wild-type COP1 transcript. The presence of small quantities of wild-type COP1 transcripts may reduce the severity of the phenotype produced by the mutated COP1' protein. The genomic DNA sequences of the COP1 gene from wild-type and lip1 peas and the cDNA sequences of COP1 and COP1' transcripts have been submitted to the EMBL database under the EMBL accession numbers AJ276591, AJ276592, AJ289773, and AJ289774, respectively.
Figures
Similar articles
-
The distribution of protochlorophyllide and chlorophyll within seedlings of the lip1 mutant of Pea.Plant Cell Physiol. 2001 Sep;42(9):931-41. doi: 10.1093/pcp/pce118. Plant Cell Physiol. 2001. PMID: 11577187
-
Molecular cloning and sequencing of the cDNA of cop1 gene from Pisum sativum.Biochim Biophys Acta. 1998 Feb 11;1395(3):326-8. doi: 10.1016/s0167-4781(97)00200-5. Biochim Biophys Acta. 1998. PMID: 9512668
-
COP1 plays a prominent role in drought stress tolerance in Arabidopsis and Pea.Plant Physiol Biochem. 2018 Sep;130:678-691. doi: 10.1016/j.plaphy.2018.08.015. Epub 2018 Aug 9. Plant Physiol Biochem. 2018. PMID: 30139551
-
Structural organization and interactions of COP1, a light-regulated developmental switch.Plant Mol Biol. 1999 Sep;41(2):151-8. doi: 10.1023/a:1006324115086. Plant Mol Biol. 1999. PMID: 10579483 Review.
-
The COP/DET/FUS proteins-regulators of eukaryotic growth and development.Semin Cell Dev Biol. 2000 Dec;11(6):495-503. doi: 10.1006/scdb.2000.0203. Semin Cell Dev Biol. 2000. PMID: 11145879 Review.
Cited by
-
Transcriptional regulation of human eosinophil RNases by an evolutionary- conserved sequence motif in primate genome.BMC Mol Biol. 2007 Oct 11;8:89. doi: 10.1186/1471-2199-8-89. BMC Mol Biol. 2007. PMID: 17927842 Free PMC article.
-
PEP1 of Arabis alpina is encoded by two overlapping genes that contribute to natural genetic variation in perennial flowering.PLoS Genet. 2012;8(12):e1003130. doi: 10.1371/journal.pgen.1003130. Epub 2012 Dec 20. PLoS Genet. 2012. PMID: 23284298 Free PMC article.
-
Ethylene-independent promotion of photomorphogenesis in the dark by cytokinin requires COP1 and the CDD complex.J Exp Bot. 2019 Jan 1;70(1):165-178. doi: 10.1093/jxb/ery344. J Exp Bot. 2019. PMID: 30272197 Free PMC article.
-
PAPP5 is involved in the tetrapyrrole mediated plastid signalling during chloroplast development.PLoS One. 2013;8(3):e60305. doi: 10.1371/journal.pone.0060305. Epub 2013 Mar 29. PLoS One. 2013. PMID: 23555952 Free PMC article.
-
The GmSTF1/2-GmBBX4 negative feedback loop acts downstream of blue-light photoreceptors to regulate isoflavonoid biosynthesis in soybean.Plant Commun. 2024 Feb 12;5(2):100730. doi: 10.1016/j.xplc.2023.100730. Epub 2023 Oct 10. Plant Commun. 2024. PMID: 37817409 Free PMC article.
References
-
- Ahmad, M., and Cashmore, A. (1993). HY4 gene of A. thaliana encodes a protein with characteristics of a blue light photoreceptor. Nature 360, 162–166. - PubMed
-
- Ang, L.H., Chattopadhyay, S., Wei, N., Oyama, T., Okada, K., Batschauer, A., and Deng, X.-W. (1998). Molecular interaction between COP1 and HY5 defines a regulatory switch for light control of Arabidopsis development. Mol. Cell 1, 213–222. - PubMed
-
- Bechtold, N., Ellis, J., and Pelletier, G. (1993). In planta Agrobacterium-mediated gene-transfer by infiltration of adult Arabidopsis thaliana plants. C. R. Acad. Sci. Ser. III 316, 1194–1199.
-
- Chamovitz, D.A., Wei, N., Osterlund, M.T., von Arnim, A., Staub, J.M., Matsui, M., and Deng, X.W. (1996). The COP9 complex, a novel multisubunit nuclear regulator involved in light control of a plant developmental switch. Cell 86, 115–121. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
