

Genetic interactions during root hair morphogenesis in Arabidopsis
- PMID: 11041890
- PMCID: PMC149133
- DOI: 10.1105/tpc.12.10.1961
Genetic interactions during root hair morphogenesis in Arabidopsis
Abstract
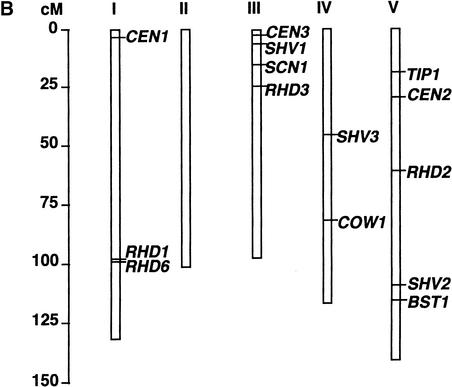
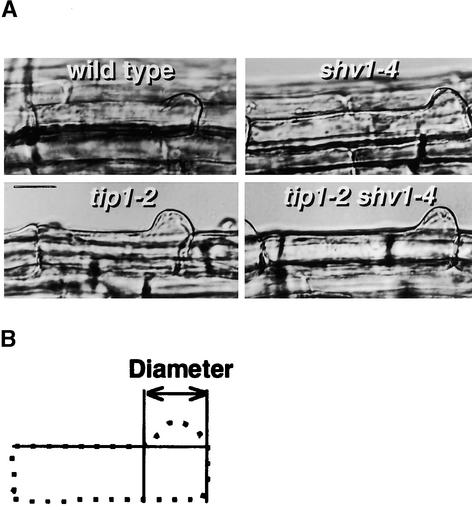
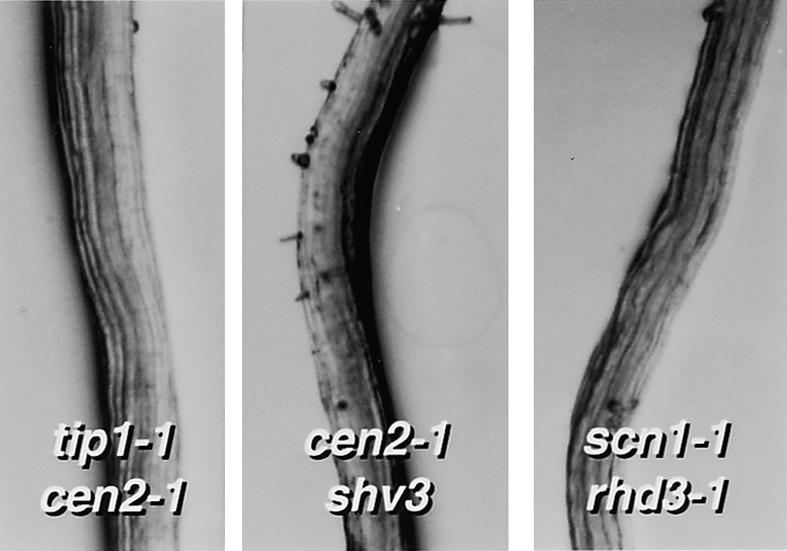
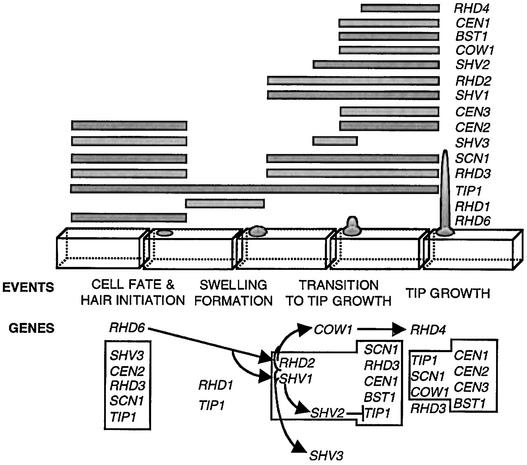
Root hairs are a major site for the uptake of water and nutrients into plants and form an increasingly important model system for studies of development of higher plants and cell biology. We have identified loss-of-function mutations in eight new genes required for hair growth in Arabidopsis: SHAVEN1 (SHV1), SHV2, and SHV3; CENTIPEDE1 (CEN1), CEN2, and CEN3; BRISTLED1 (BST1); and SUPERCENTIPEDE1 (SCN1). We combined mutations in 79 pairs of genes to determine the stages at which these and six previously known genes contribute to root hair formation. Double mutant phenotypes revealed roles for several genes that could not have been predicted from the single mutant phenotypes. For example, we show that TIP1 and RHD3 are required much earlier in hair formation than previous studies have suggested. We present a genetic model for root hair morphogenesis that defines the roles of each gene, and we suggest hypotheses about functional relationships between genes.
Figures




References
-
- Bibikova, T.N., Jacob, T., Dahse, I., and Gilroy, S. (1998). Localized changes in apoplastic and cytoplasmic pH are associated with root hair development in Arabidopsis thaliana. Development 125, 2925–2934. - PubMed
-
- DiCristina, M., Sessa, G., Dolan, L., Linstead, P., Baima, S., Ruberti, I., and Morelli, G. (1996). The Arabidopsis Athb-10 (GLABRA2) is an HD-Zip protein required for regulation of root hair development. Plant J. 10, 393–402. - PubMed
-
- Dolan, L., and Scheres, B. (1998). Root pattern: Shooting in the dark? Semin. Cell Dev. Biol. 9, 201–206. - PubMed
-
- Dolan, L., Duckett, C.M., Grierson, C., Linstead, P., Schneider, K., Lawson, E., Dean, C., Poethig, S., and Roberts, K. (1994). Clonal relationships and cell patterning in the root epidermis of Arabidopsis. Development 120, 2465–2474.
-
- Galway, M.E., Masucci, J.D., Lloyd, A.M., Walbot, V., Davis, R.W., and Schiefelbein, J.W. (1994). The TTG gene is required to specify epidermal-cell fate and cell patterning in the Arabidopsis root. Dev. Biol. 166, 740–754. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
