

Maternal environment and genotype interact to establish diabesity in mice
- PMID: 11042154
- PMCID: PMC310941
- DOI: 10.1101/gr.147000
Maternal environment and genotype interact to establish diabesity in mice
Abstract
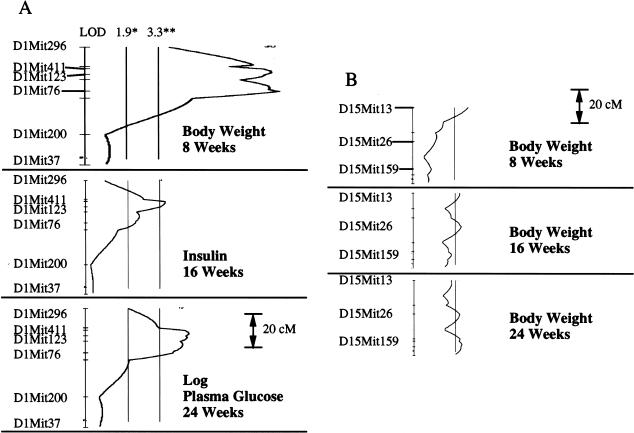
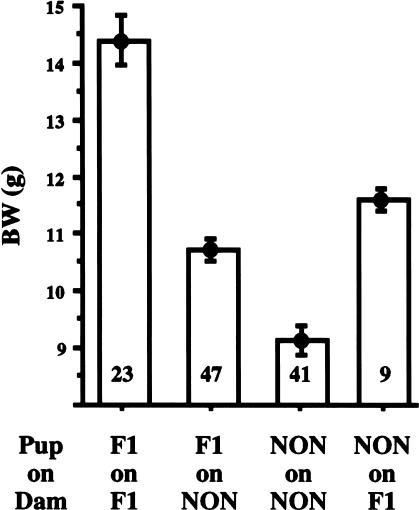
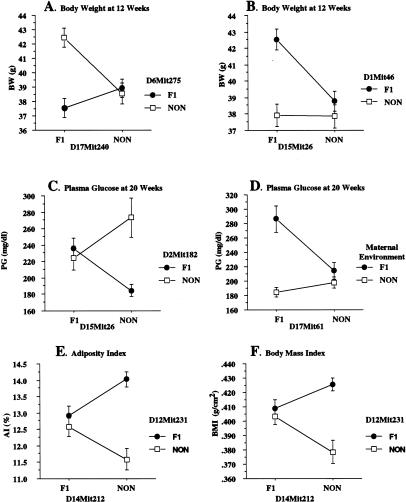
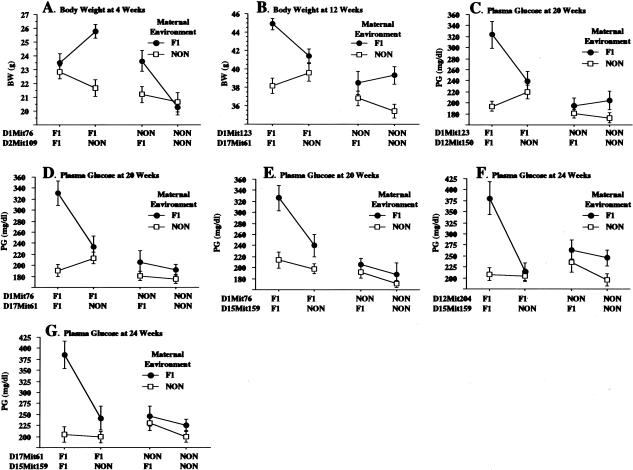
Obesity, a major risk factor for type II diabetes, is becoming more prevalent in Western populations consuming high calorie diets while expending less energy both at the workplace and at home. Most human obesity, and probably most type II diabetes as well, reflects polygenic rather than monogenic inheritance. We have genetically dissected a polygenic mouse model of obesity-driven type II diabetes by outcrossing the obese, diabetes-prone, NZO (New Zealand Obese)/HlLt strain to the relatively lean NON (Nonobese Nondiabetic)/Lt strain, and then reciprocally backcrossing obese F1 mice to the lean NON/Lt parental strain. A continuous distribution of body weights was observed in a population of 203 first backcross males. The 22% of first backcross males developing overt diabetes showed highest peripubertal weight gains and earliest development of hyperinsulinemia. We report a complex diabetes-predisposing ("diabesity") QTL (Quantitative Trait Loci) on chromosome 1 contributing significant main effects to increases in body weight, plasma insulin, and plasma glucose. NZO contributed QTL with significant main effects on adiposity parameters on chromosomes 12 and 5. A NON QTL on chromosome 14 interacted epistatically with the NZO obesity QTL on chromosome 12 to increase adiposity. Although the main effect of the diabetogenic QTL on chromosome 1 was on rapid growth rather than adiposity, it interacted epistatically with the obesity QTL on chromosome 12 to increase plasma glucose levels. Additional complex epistatic interactions eliciting significant increases in body weight and/or plasma glucose were found between the NZO-contributed QTL on chromosome 1 and other NZO-contributed QTL on chromosomes 15 and 17, as well as with an NON-contributed QTL on chromosome 2. We further show that certain of these intergenic interactions are predicated on, or enhanced by, the maternal postparturitional environment. We show by cross-fostering experiments that the maternal environmental influence in part is because of the presence of early obesity-inducing factors in the milk of obese F1 dams. We also discuss a strategy for using recombinant congenic strains to separate and reassemble interacting QTL for future study.
Figures

Similar articles
-
Deconstructing and reconstructing obesity-induced diabetes (diabesity) in mice.Diabetes. 2002 Mar;51(3):825-32. doi: 10.2337/diabetes.51.3.825. Diabetes. 2002. PMID: 11872687
-
Differential levels of diabetogenic stress in two new mouse models of obesity and type 2 diabetes.Diabetes. 2004 Feb;53 Suppl 1:S4-11. doi: 10.2337/diabetes.53.2007.s4. Diabetes. 2004. PMID: 14749259 Review.
-
Characterisation of the mouse diabetes susceptibilty locus Nidd/SJL: islet cell destruction, interaction with the obesity QTL Nob1, and effect of dietary fat.Diabetologia. 2002 Jun;45(6):823-30. doi: 10.1007/s00125-002-0796-7. Epub 2002 Apr 26. Diabetologia. 2002. PMID: 12107726
-
Characterization of Nob3, a major quantitative trait locus for obesity and hyperglycemia on mouse chromosome 1.Physiol Genomics. 2009 Jul 9;38(2):226-32. doi: 10.1152/physiolgenomics.00011.2009. Epub 2009 May 26. Physiol Genomics. 2009. PMID: 19470805 Free PMC article.
-
The genetic basis of obesity and type 2 diabetes: lessons from the new zealand obese mouse, a polygenic model of the metabolic syndrome.Results Probl Cell Differ. 2010;52:1-11. doi: 10.1007/978-3-642-14426-4_1. Results Probl Cell Differ. 2010. PMID: 20865367 Review.
Cited by
-
'Fat's chances': Loci for phenotypic dispersion in plasma leptin in mouse models of diabetes mellitus.PLoS One. 2019 Oct 29;14(10):e0222654. doi: 10.1371/journal.pone.0222654. eCollection 2019. PLoS One. 2019. PMID: 31661517 Free PMC article.
-
Bioactive compounds in mothers milk affecting offspring outcomes: A narrative review.Pediatr Obes. 2022 Jul;17(7):e12892. doi: 10.1111/ijpo.12892. Epub 2022 Jan 20. Pediatr Obes. 2022. PMID: 35060344 Free PMC article. Review.
-
Effects of early postnatal environment on hypothalamic gene expression in OLETF rats.PLoS One. 2017 Jun 2;12(6):e0178428. doi: 10.1371/journal.pone.0178428. eCollection 2017. PLoS One. 2017. PMID: 28575002 Free PMC article.
-
Lineage is an epigenetic modifier of QTL influencing behavioral coping with stress.Behav Genet. 2005 Mar;35(2):189-98. doi: 10.1007/s10519-004-1018-5. Behav Genet. 2005. PMID: 15685431 Free PMC article.
-
Fostering in mice induces cardiovascular and metabolic dysfunction in adulthood.J Physiol. 2011 Aug 15;589(Pt 16):3969-81. doi: 10.1113/jphysiol.2011.212324. Epub 2011 Jun 13. J Physiol. 2011. PMID: 21669973 Free PMC article.
References
-
- Bjorntorp P. Androgens, the metabolic syndrome, and non-insulin dependent diabetes mellitus. Ann NY Acad Sci. 1993;676:242–252. - PubMed
-
- Clark AG. Limits to prediction of phenotype from knowledge of genotypes. In: Clegg MT, Hecht M, MacIntyre RJ, editors. Limits to knowledge in evolutionary genetics. New York: Kluwer Academic/Plenum Publishers; 2000. pp. 205–224.
-
- Cox NJ, Frigge M, Nicolae DL, Concannon P, Hanis CL, Bell GI, Kong A. Loci on chromosomes 2 (NIDDM1) and 15 interact to increase susceptibility to diabetes in Mexican Americans. Nat Genet. 1999;21:213–215. - PubMed
-
- Flurkey K, Rosen C, Partke H-J, Herberg L, Leiter EH. Rapid maturational growth as a predictor of NIDDM in NZO male mice. Diabetes. 1998;47: (Suppl.):A318. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases