

Selection of optimal polypurine tract region sequences during Moloney murine leukemia virus replication
- PMID: 11044073
- PMCID: PMC110903
- DOI: 10.1128/jvi.74.22.10293-10303.2000
Selection of optimal polypurine tract region sequences during Moloney murine leukemia virus replication
Abstract
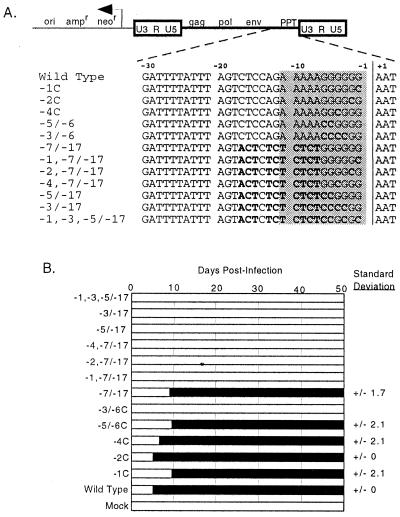
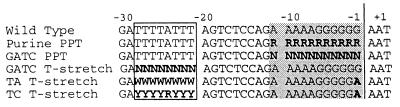
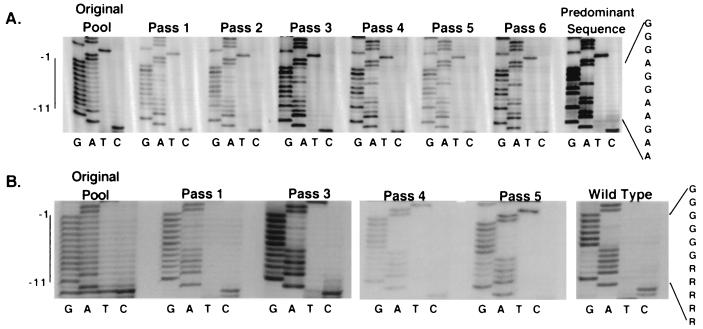
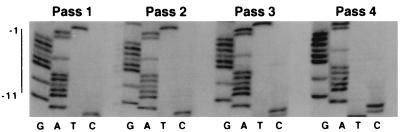
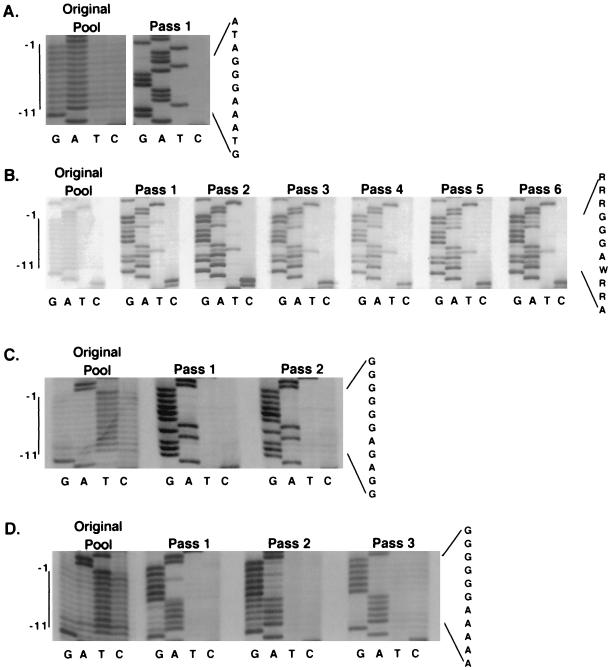
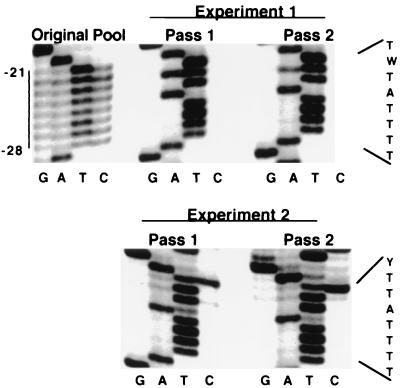
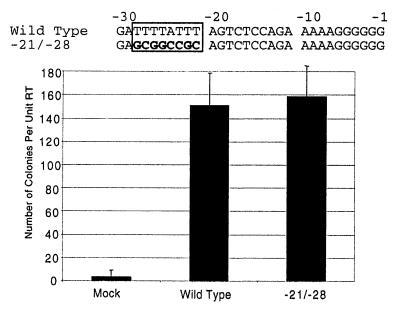
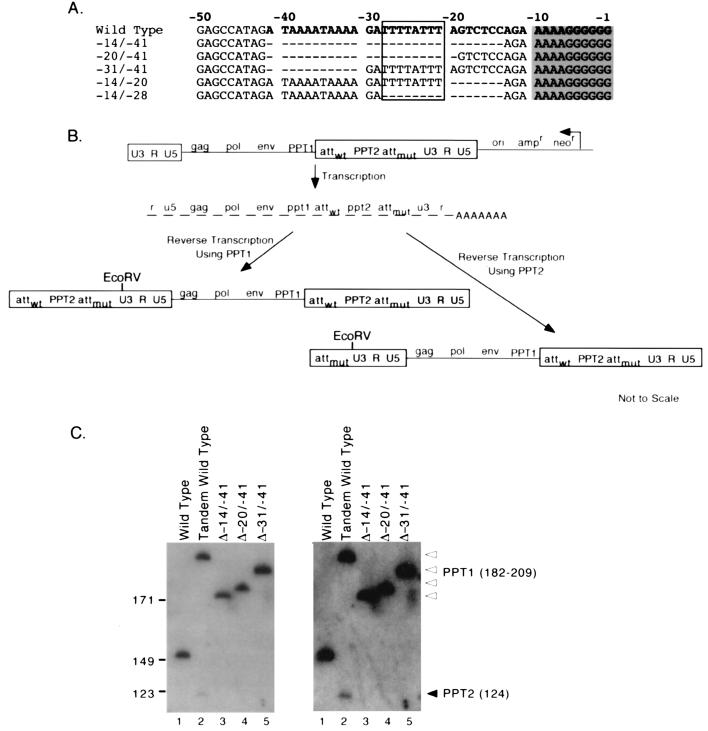
Retrovirus plus-strand synthesis is primed by a cleavage remnant of the polypurine tract (PPT) region of viral RNA. In this study, we tested replication properties for Moloney murine leukemia viruses with targeted mutations in the PPT and in conserved sequences upstream, as well as for pools of mutants with randomized sequences in these regions. The importance of maintaining some purine residues within the PPT was indicated both by examining the evolution of random PPT pools and from the replication properties of targeted mutants. Although many different PPT sequences could support efficient replication and one mutant that contained two differences in the core PPT was found to replicate as well as the wild type, some sequences in the core PPT clearly conferred advantages over others. Contributions of sequences upstream of the core PPT were examined with deletion mutants. A conserved T-stretch within the upstream sequence was examined in detail and found to be unimportant to helper functions. Evolution of virus pools containing randomized T-stretch sequences demonstrated marked preference for the wild-type sequence in six of its eight positions. These findings demonstrate that maintenance of the T-rich element is more important to viral replication than is maintenance of the core PPT.
Figures
References
-
- Champoux J J. Roles of ribonuclease H in reverse transcription. In: Skalka A M, Goff S P, editors. Reverse transcriptase. Cold Spring Harbor, N.Y: Cold Spring Harbor Laboratory; 1993. pp. 103–117.
-
- Fedoroff O Y, Ge Y, Reid B R. Solution structure of r(gaggacug):d(CAGTCCTC) hybrid: implications for the initiation of HIV-1 (+)-strand synthesis. J Mol Biol. 1997;269:225–239. - PubMed
-
- Gao H-Q, Boyer P L, Arnold E, Hughes S H. Effects of mutations in the polymerase domain on the polymerase, RNase H and strand transfer activities of human immunodeficiency virus type 1 reverse transcriptase. J Mol Biol. 1998;277:559–572. - PubMed
-
- Gao H-Q, Sarafianos S G, Arnold E, Hughes S H. Similarities and differences in the RNaseH activities of human immunodeficiecy virus type 1 reverse transcriptase and Moloney murine leukemia virus reverse transcriptase. J Mol Biol. 1999;294:1097–1113. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
