

Utilization of the bovine papillomavirus type 1 late-stage-specific nucleotide 3605 3' splice site is modulated by a novel exonic bipartite regulator but not by an intronic purine-rich element
- PMID: 11044105
- PMCID: PMC110935
- DOI: 10.1128/jvi.74.22.10612-10622.2000
Utilization of the bovine papillomavirus type 1 late-stage-specific nucleotide 3605 3' splice site is modulated by a novel exonic bipartite regulator but not by an intronic purine-rich element
Abstract
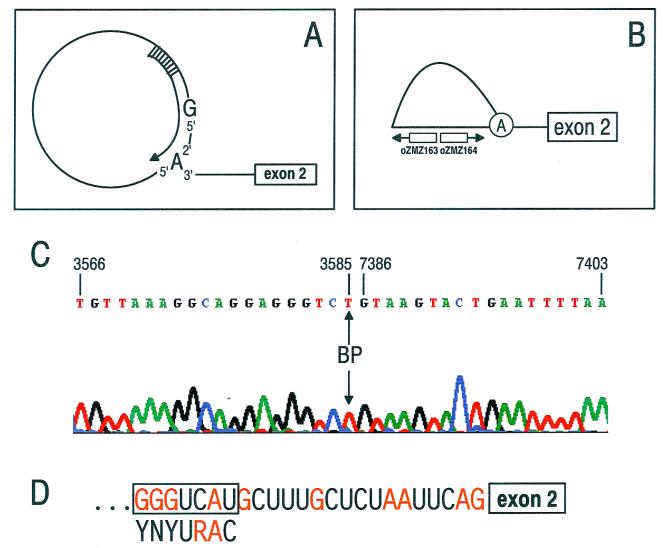
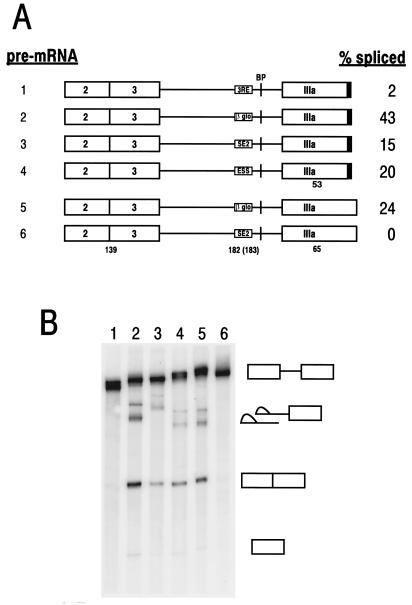
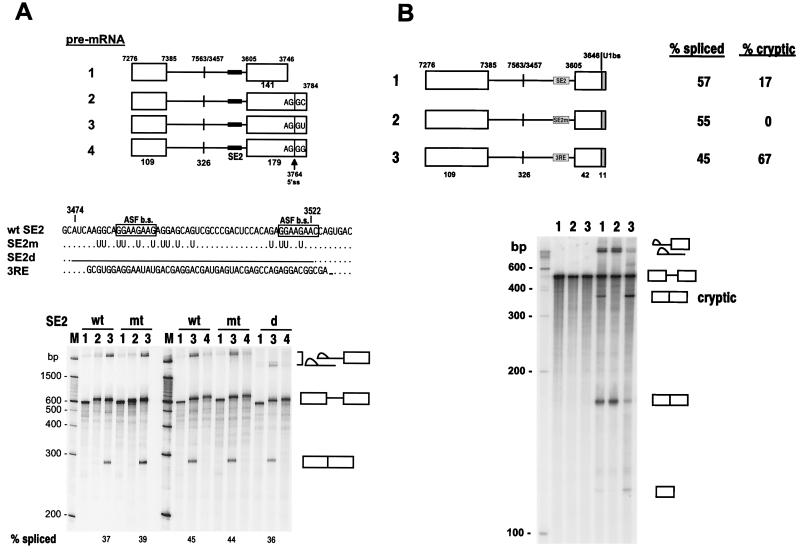
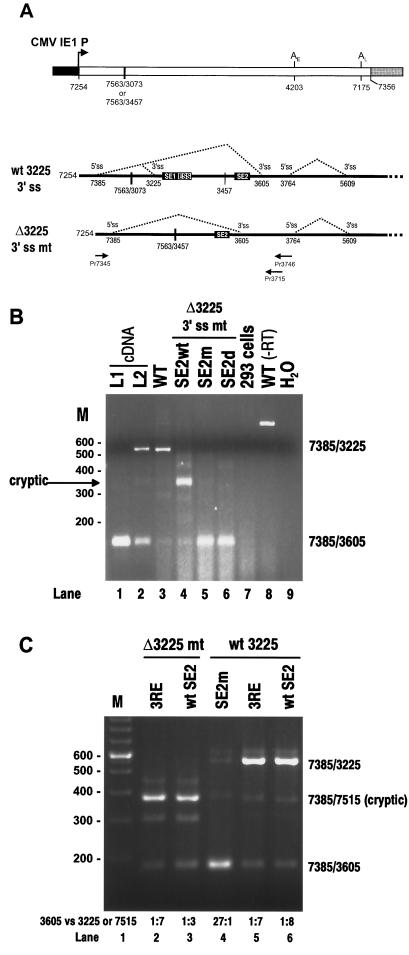
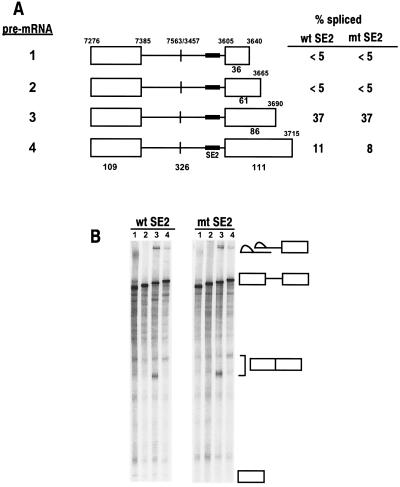
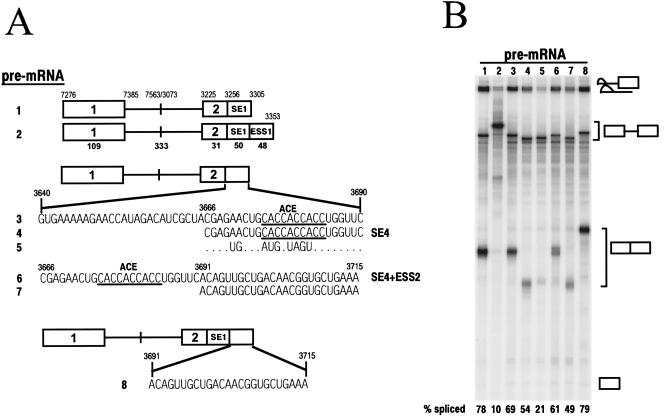
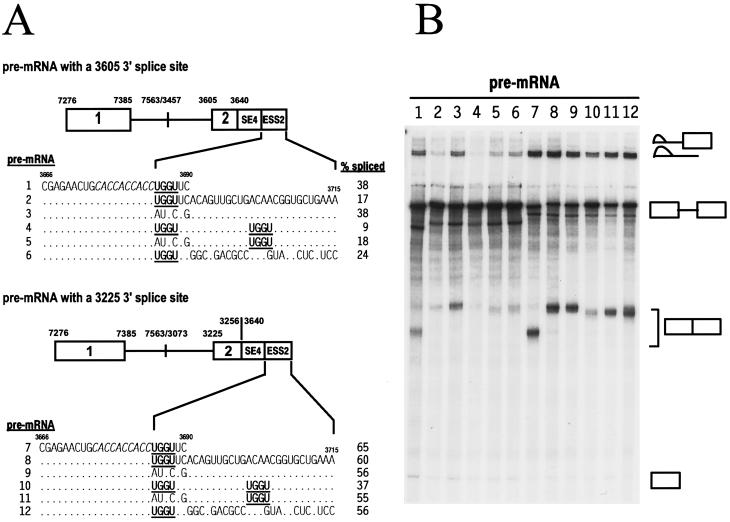
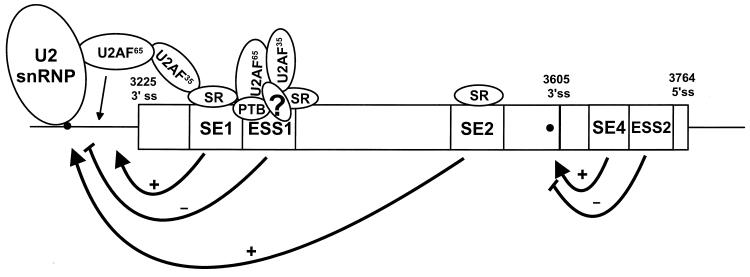
Bovine papillomavirus type 1 (BPV-1) late gene expression is regulated at both transcriptional and posttranscriptional levels. Maturation of the capsid protein (L1) pre-mRNA requires a switch in 3' splice site utilization. This switch involves activation of the nucleotide (nt) 3605 3' splice site, which is utilized only in fully differentiated keratinocytes during late stages of the virus life cycle. Our previous studies of the mechanisms that regulate BPV-1 alternative splicing identified three cis-acting elements between these two splice sites. Two purine-rich exonic splicing enhancers, SE1 and SE2, are essential for preferential utilization of the nt 3225 3' splice site at early stages of the virus life cycle. Another cis-acting element, exonic splicing suppressor 1 (ESS1), represses use of the nt 3225 3' splice site. In the present study, we investigated the late-stage-specific nt 3605 3' splice site and showed that it has suboptimal features characterized by a nonconsensus branch point sequence and a weak polypyrimidine track with interspersed purines. In vitro and in vivo experiments showed that utilization of the nt 3605 3' splice site was not affected by SE2, which is intronically located with respect to the nt 3605 3' splice site. The intronic location and sequence composition of SE2 are similar to those of the adenovirus IIIa repressor element, which has been shown to inhibit use of a downstream 3' splice site. Further studies demonstrated that the nt 3605 3' splice site is controlled by a novel exonic bipartite element consisting of an AC-rich exonic splicing enhancer (SE4) and an exonic splicing suppressor (ESS2) with a UGGU motif. Functionally, this newly identified bipartite element resembles the bipartite element composed of SE1 and ESS1. SE4 also functions on a heterologous 3' splice site. In contrast, ESS2 functions as an exonic splicing suppressor only in a 3'-splice-site-specific and enhancer-specific manner. Our data indicate that BPV-1 splicing regulation is very complex and is likely to be controlled by multiple splicing factors during keratinocyte differentiation.
Figures
Similar articles
-
Selection of the bovine papillomavirus type 1 nucleotide 3225 3' splice site is regulated through an exonic splicing enhancer and its juxtaposed exonic splicing suppressor.J Virol. 1996 Jul;70(7):4691-9. doi: 10.1128/JVI.70.7.4691-4699.1996. J Virol. 1996. PMID: 8676495 Free PMC article.
-
Optimization of a weak 3' splice site counteracts the function of a bovine papillomavirus type 1 exonic splicing suppressor in vitro and in vivo.J Virol. 2000 Jul;74(13):5902-10. doi: 10.1128/jvi.74.13.5902-5910.2000. J Virol. 2000. PMID: 10846071 Free PMC article.
-
Function of a bovine papillomavirus type 1 exonic splicing suppressor requires a suboptimal upstream 3' splice site.J Virol. 1999 Jan;73(1):29-36. doi: 10.1128/JVI.73.1.29-36.1999. J Virol. 1999. PMID: 9847303 Free PMC article.
-
Alterations of pre-mRNA splicing in cancer.Genes Chromosomes Cancer. 2005 Apr;42(4):342-57. doi: 10.1002/gcc.20156. Genes Chromosomes Cancer. 2005. PMID: 15648050 Review.
-
Regulation of alternative RNA splicing by exon definition and exon sequences in viral and mammalian gene expression.J Biomed Sci. 2004 May-Jun;11(3):278-94. doi: 10.1007/BF02254432. J Biomed Sci. 2004. PMID: 15067211 Free PMC article. Review.
Cited by
-
Exonic splicing enhancer-dependent selection of the bovine papillomavirus type 1 nucleotide 3225 3' splice site can be rescued in a cell lacking splicing factor ASF/SF2 through activation of the phosphatidylinositol 3-kinase/Akt pathway.J Virol. 2003 Feb;77(3):2105-15. doi: 10.1128/jvi.77.3.2105-2115.2003. J Virol. 2003. PMID: 12525645 Free PMC article.
-
A splicing enhancer in the E4 coding region of human papillomavirus type 16 is required for early mRNA splicing and polyadenylation as well as inhibition of premature late gene expression.J Virol. 2005 Sep;79(18):12002-15. doi: 10.1128/JVI.79.18.12002-12015.2005. J Virol. 2005. PMID: 16140776 Free PMC article.
-
Systematic characterization of short intronic splicing-regulatory elements in SMN2 pre-mRNA.Nucleic Acids Res. 2022 Jan 25;50(2):731-749. doi: 10.1093/nar/gkab1280. Nucleic Acids Res. 2022. PMID: 35018432 Free PMC article.
-
Kaposi's sarcoma-associated herpesvirus K8beta is derived from a spliced intermediate of K8 pre-mRNA and antagonizes K8alpha (K-bZIP) to induce p21 and p53 and blocks K8alpha-CDK2 interaction.J Virol. 2005 Nov;79(22):14207-21. doi: 10.1128/JVI.79.22.14207-14221.2005. J Virol. 2005. PMID: 16254356 Free PMC article.
-
Multiple regions of Kaposi's sarcoma-associated herpesvirus ORF59 RNA are required for its expression mediated by viral ORF57 and cellular RBM15.Viruses. 2015 Feb 3;7(2):496-510. doi: 10.3390/v7020496. Viruses. 2015. PMID: 25690794 Free PMC article.
References
-
- Adams M D, Rudner D Z, Rio D C. Biochemistry and regulation of pre-mRNA splicing. Curr Opin Cell Biol. 1996;8:331–339. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
