

Multiple C-terminal lysine residues target p53 for ubiquitin-proteasome-mediated degradation
- PMID: 11046142
- PMCID: PMC102152
- DOI: 10.1128/MCB.20.22.8458-8467.2000
Multiple C-terminal lysine residues target p53 for ubiquitin-proteasome-mediated degradation
Abstract
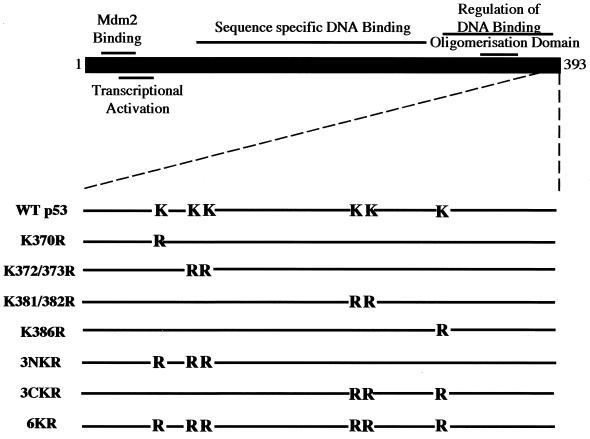
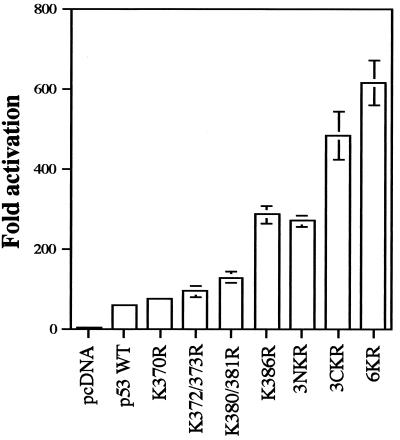
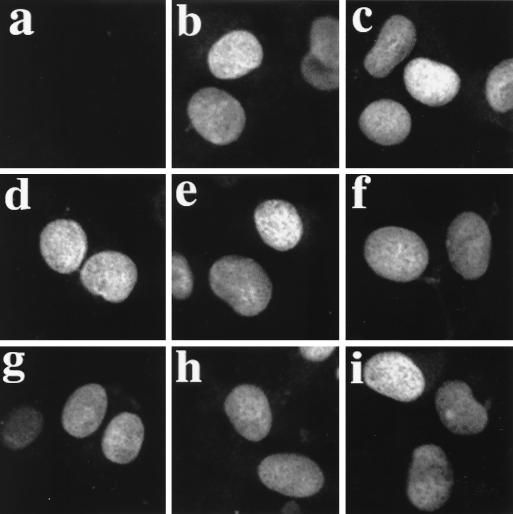
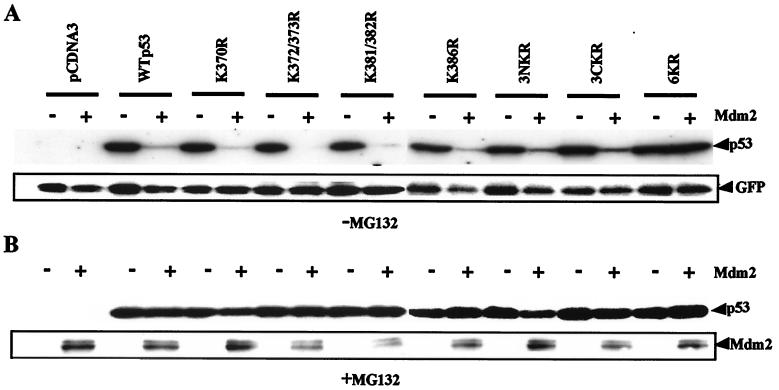
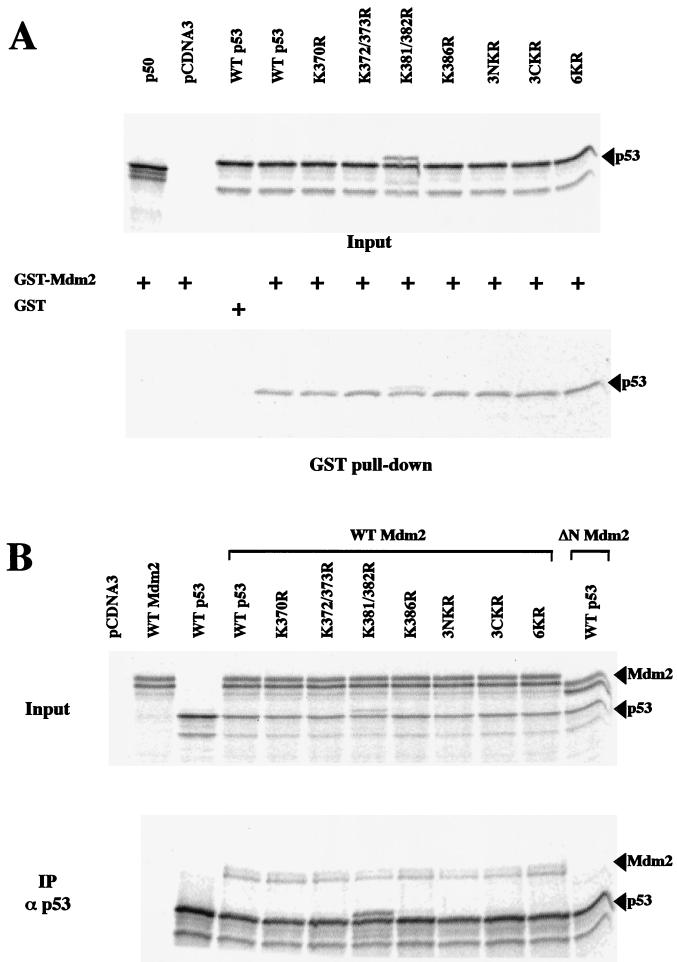
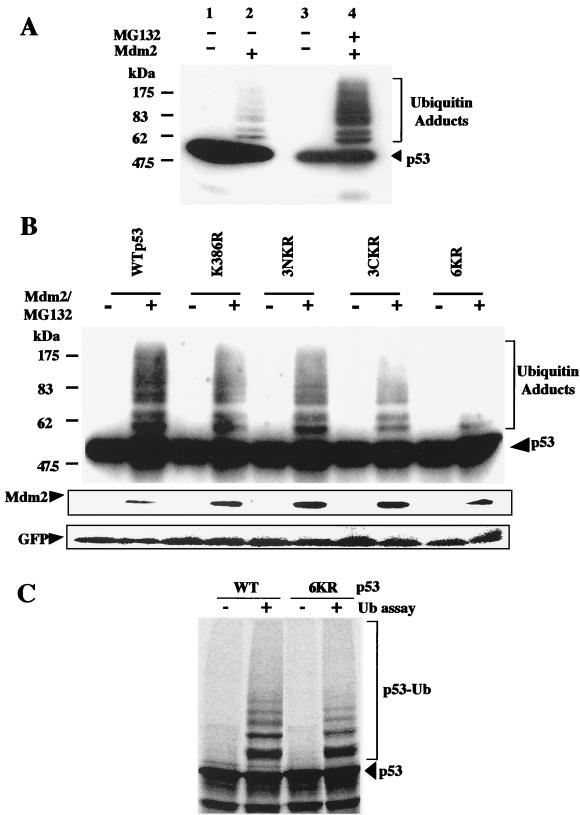
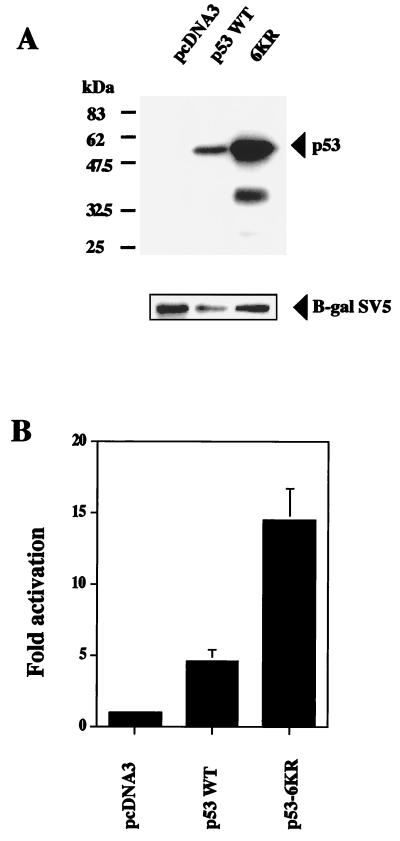
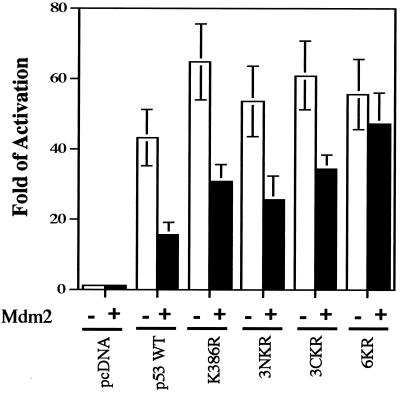
In normal cells, p53 is maintained at a low level by ubiquitin-mediated proteolysis, but after genotoxic insult this process is inhibited and p53 levels rise dramatically. Ubiquitination of p53 requires the ubiquitin-activating enzyme Ubc5 as a ubiquitin conjugation enzyme and Mdm2, which acts as a ubiquitin protein ligase. In addition to the N-terminal region, which is required for interaction with Mdm2, the C-terminal domain of p53 modulates the susceptibility of p53 to Mdm2-mediated degradation. To analyze the role of the C-terminal domain in p53 ubiquitination, we have generated p53 molecules containing single and multiple lysine-to-arginine changes between residues 370 and 386. Although wild-type (WT) and mutant molecules show similar subcellular distributions, the mutants display a higher transcriptional activity than WT p53. Simultaneous mutation of lysine residues 370, 372, 373, 381, 382, and 386 to arginine residues (6KR p53 mutant) generates a p53 molecule with potent transcriptional activity that is resistant to Mdm2-induced degradation and is refractory to Mdm2-mediated ubiquitination. In contrast to WT p53, transcriptional activity directed by the 6KR p53 mutant fails to be negatively regulated by Mdm2. Those differences are also manifest in HeLa cells which express the human papillomavirus E6 protein, suggesting that p53 C-terminal lysine residues are also implicated in E6-AP-mediated ubiquitination. These data suggest that p53 C-terminal lysine residues are the main sites of ubiquitin ligation, which target p53 for proteasome-mediated degradation.
Figures
References
-
- Askjaer P, Jensen T H, Nilsson J, Englmeier L, Kjems J. The specificity of the CRM1-Rev nuclear export signal interaction is mediated by RanGTP. J Biol Chem. 1998;273:33414–33422. - PubMed
-
- Baldi L, Brown K, Franzoso G, Siebenlist U. Critical role for lysine-21 and lysine-22 signal-induced, ubiquitin-mediated proteolysis of IκBα. J Biol Chem. 1996;271:376–379. - PubMed
-
- Bottger V, Bottger A, Howard S F, Picksley S M, Chene P, Garcia-Echeverria C, Hochkeppel H K, Lane D P. Identification of novel mdm2 binding peptides by phage display. Oncogene. 1996;13:2141–2147. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous