

The hsp110 and Grp1 70 stress proteins: newly recognized relatives of the Hsp70s
- PMID: 11048651
- PMCID: PMC312858
- DOI: 10.1379/1466-1268(2000)005<0276:thagsp>2.0.co;2
The hsp110 and Grp1 70 stress proteins: newly recognized relatives of the Hsp70s
Abstract
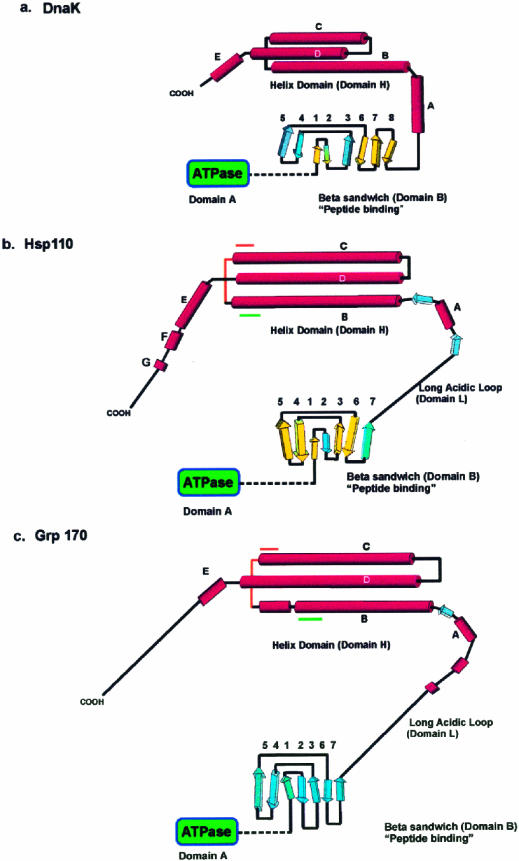
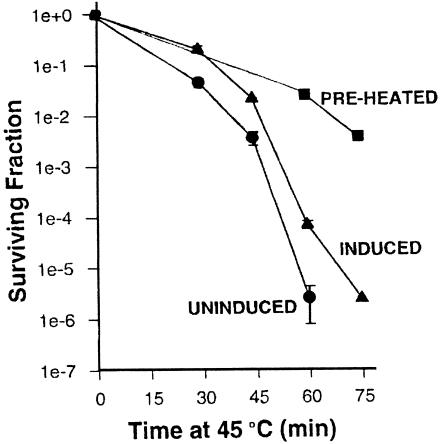
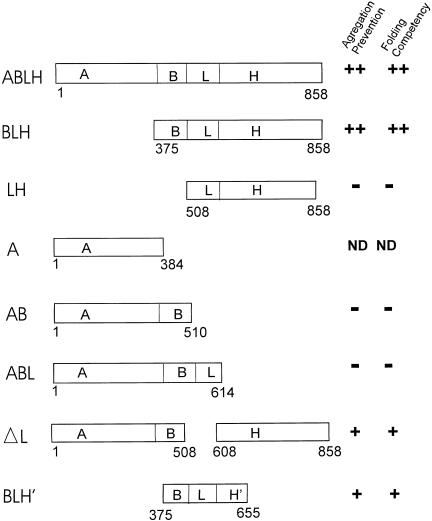
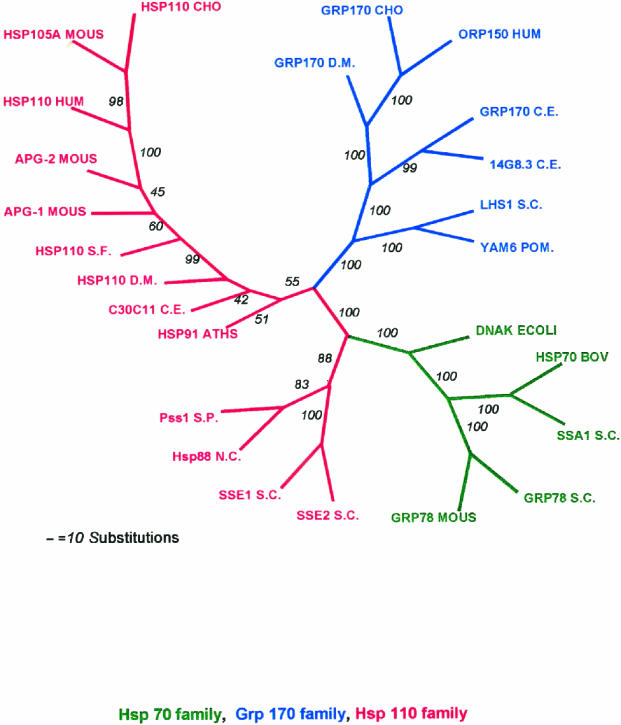
Both the Grp170 and Hsp110 families represent relatively conserved and distinct sets of stress proteins, within a more diverse category that also includes the Hsp70s. All of these families are found in a wide variety of organisms from yeasts to humans. Although Hsp110s or Grp170s are not Hsp70s any more than Hsp70s are Hsp110s or Grp170s, it is still reasonable to refer to this combination of related families as the Hsp70 superfamily based on arguments discussed above and since no obvious prokaryotic Hsp110 or Grp170 has yet been identified. These proteins are related to their counterparts in the Hsp70/Grp78 family of eukaryotic stress proteins but are characterized by significantly larger molecular weights. The members of the Grp170 family are characterized by C-terminal ER retention sequences and are ER localized in yeasts and mammals. As a Grp, Grp170 is recognized to be coregulated with other major Grps by a well-known set of stress conditions, sometimes referred to as the unfolded protein response (Kozutsumi et al 1988; Nakaki et al 1989). The Hsp110 family members are localized in the nucleus and cytoplasm and, with other major Hsps, are also coregulated by a specific set of stress conditions, most notably including hyperthermic exposures. Hsp110 is sometimes called Hsp105, although it would be preferable to have a uniform term. The large Hsp70-like proteins are structurally similar to the Hsp70s but differ from them in important ways. In both the Grp170 and Hspl10 families, there is a long loop structure that is interposed between the peptide-binding ,-domain and the alpha-helical lid. In the Hsp110 family and Grp170, there are differing degrees of expansion in the alpha-helical domain and the addition of a C-terminal loop. This gives the appearance of much larger lid domains for Hsp110 and Grp170 compared with Hsp70. Both Hsp110 and Grp170 families have relatively conserved short sequences in the alpha-helical domain in the lid, which are conserved motifs in numerous proteins (we termed these motifs Magic and TedWylee as discussed earlier). The structural differences detailed in this review result in functional differences between the large (Grp170 and Hspl10) members of the Hsp70 superfamily, the most distinctive being an increased ability of these proteins to bind (hold) denatured polypeptides compared with Hsc70, perhaps related to the enlarged C-terminal helical domain. However, there is also a major difference between these large stress proteins; Hsp110 does not bind ATP in vitro, whereas Grp170 binds ATP avidly. The role of the Grp170 and Hsp110 stress proteins in cellular physiology is not well understood. Overexpression of Hsp110 in cultured mammalian cells increases thermal tolerance. Grp170 binds to secreted proteins in the ER and may be cooperatively involved in folding these proteins appropriately. These roles are similar to those of the Hsp70 family members, and, therefore, the question arises as to the differential roles played by the larger members of the superfamily. We have discussed evidence that the large members of the superfamily cooperate with members of the Hsp70 family, and these chaperones probably interact with a large number of chaperones and cochaperones in their functional activities. The fundamental point is that Hsp110 is found in conjunction with Hsp70 in the cytoplasm (and nucleus) and Grp170 is found in conjunction with78 in tha ER in every eucaryotic cell examined from yeast to humans. This would strongly argue that Hsp110 Grp170 exhibit functions in eucaryotes not effectively performed by Hsp70s or Grp78, respectively. Of interest in this respect is the observation that all Hsp110s loss of function or deletion mutants listed in the Drosophila deletion project database are lethal. The important task for the future is to determine the roles these conserved molecular chaperones play in normal and physiologically stressed cells.
Figures
References
-
- Albukrek D, Bakon M, Moran DS, Faibel M, Epstein Y. Heat-stroke-induced cerebellar atrophy: clinical course, CT and MRI findings. Neuroradiology. 1997;39:195–197. - PubMed
-
- Becker J, Craig EA. Heat-shock proteins as molecular chaperones. Eur J Biochem. 1994;219:11–23. - PubMed
-
- Boorstein WR, Ziegelhoffer T, Craig EA. Molecular evolution of the HSP70 multigene family. J Mol Evol. 1994;38:1–17. - PubMed
-
- Brodsky JL, Werner ED, Dubas ME, Goecker JL, Kruse KB, Mc Cracken AA. The requirement for molecular chaperones during endoplasmic reticulum-associated protein degradation demonstrates that protein export and import are mechanistically distinct. J Biol Chem. 1999;274:3453–3460. - PubMed
-
- Cai JW, Henderson BW, Shen JW, Subjeck JR. Induction of glucose regulated proteins during growth of a murine tumor. J Cell Physiol. 1993;154:229–237. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous