

Visual responses in monkey areas V1 and V2 to three-dimensional surface configurations
- PMID: 11050142
- PMCID: PMC6772714
- DOI: 10.1523/JNEUROSCI.20-21-08188.2000
Visual responses in monkey areas V1 and V2 to three-dimensional surface configurations
Abstract
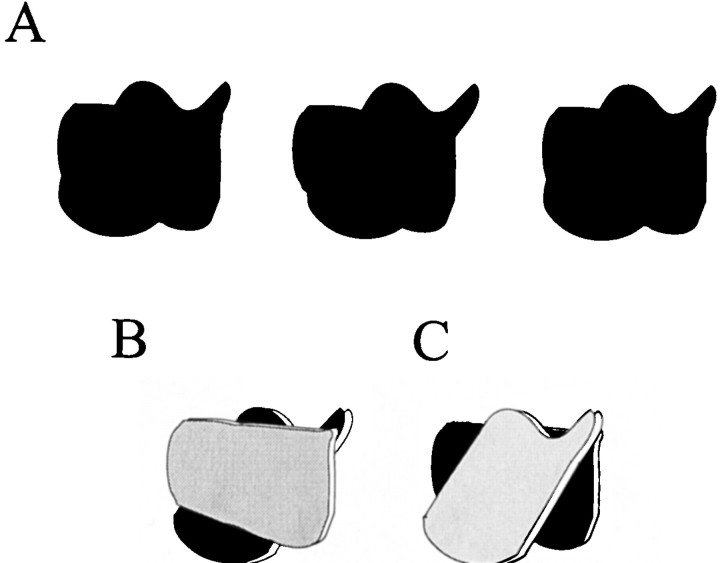
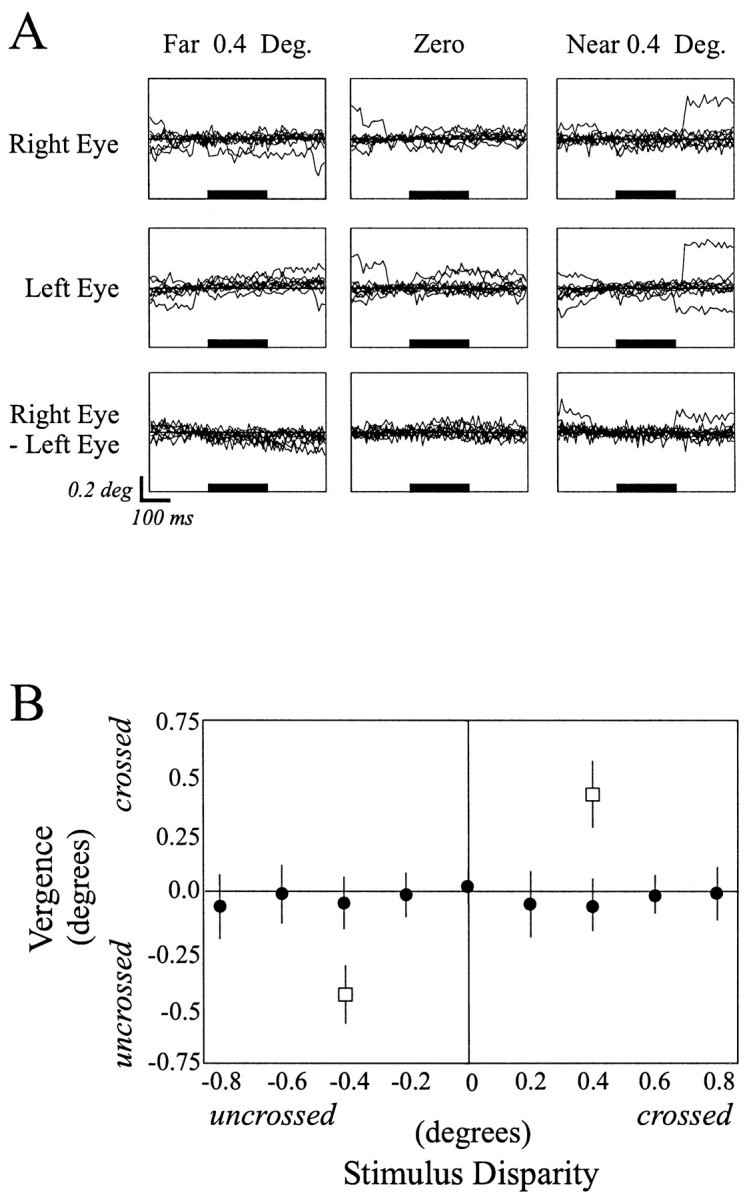
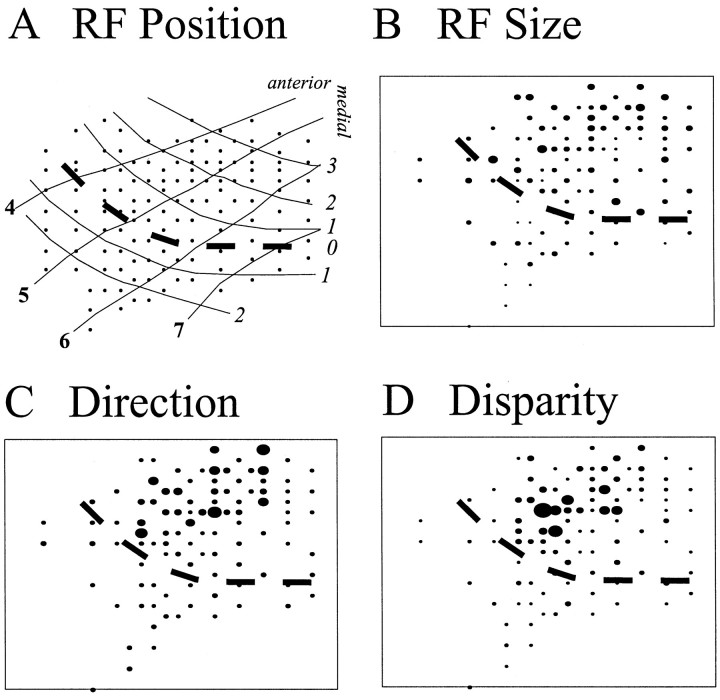
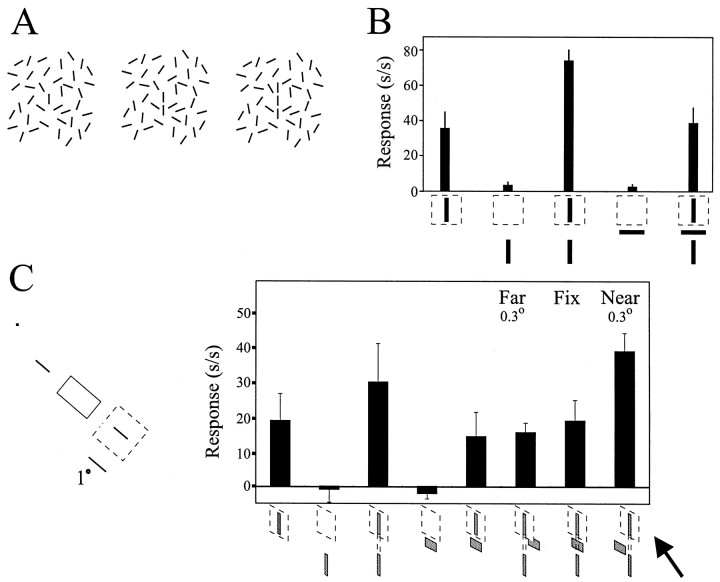
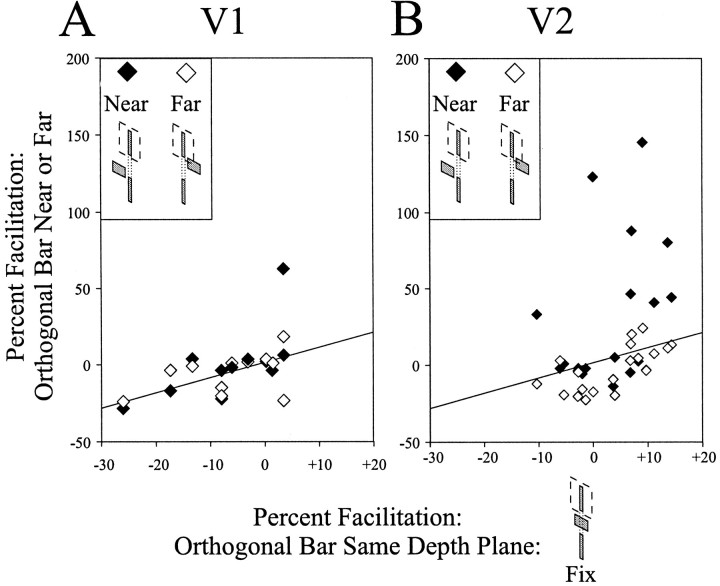
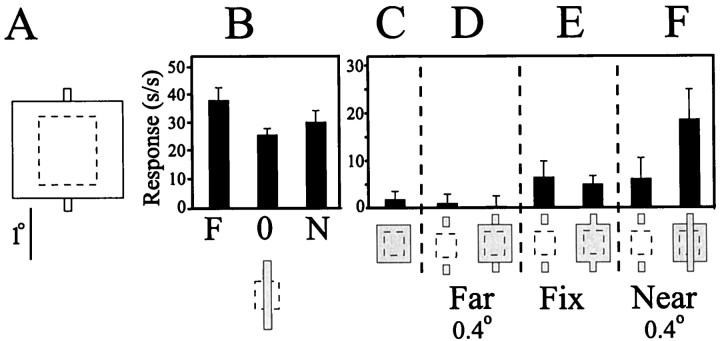
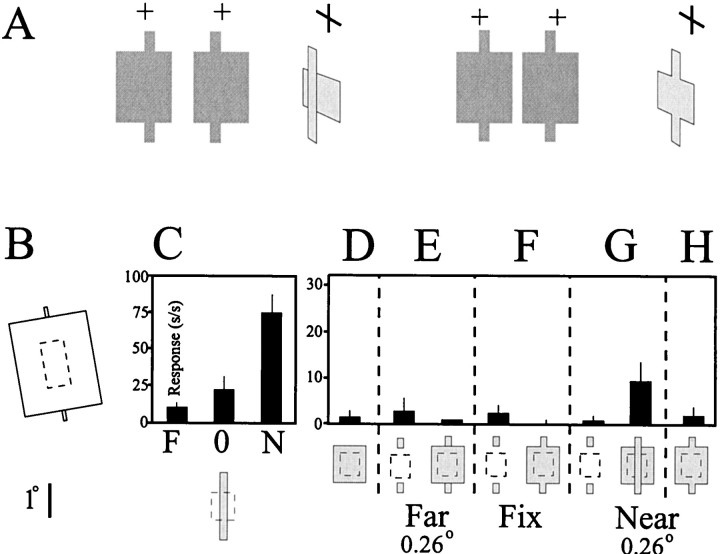
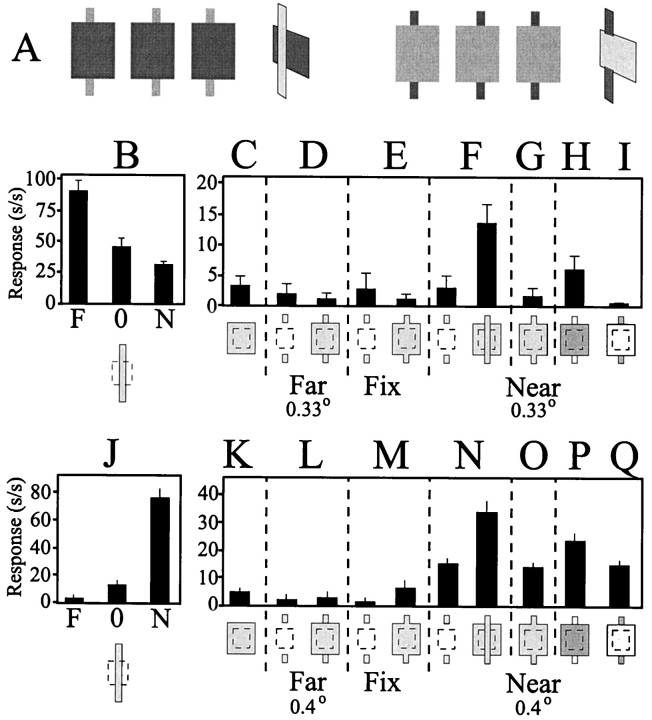
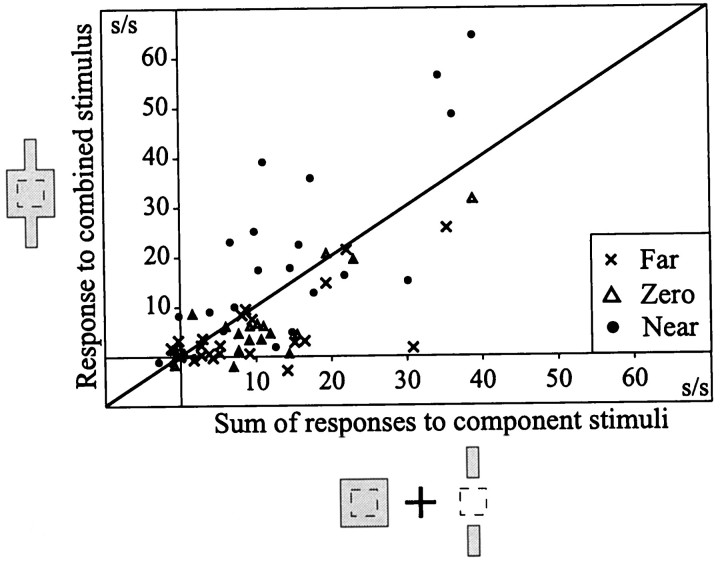
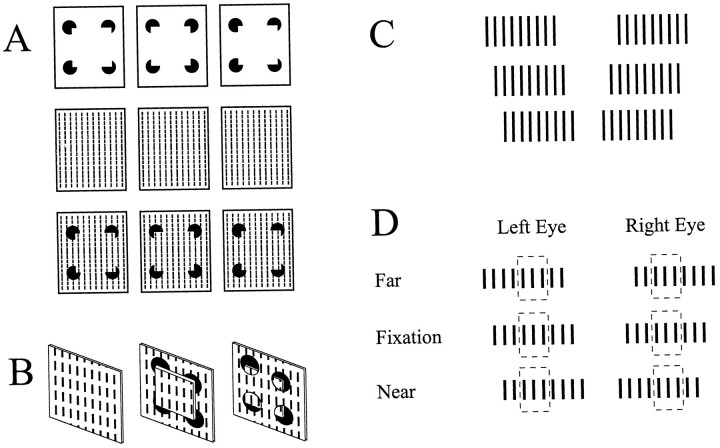
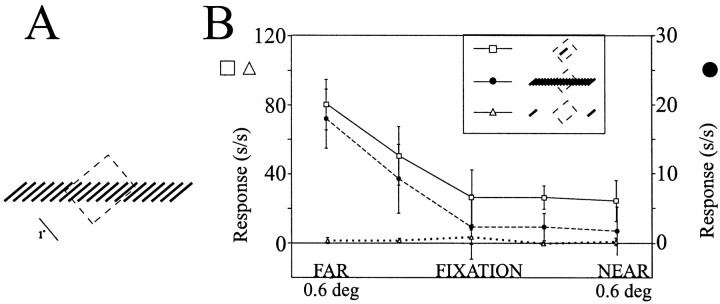
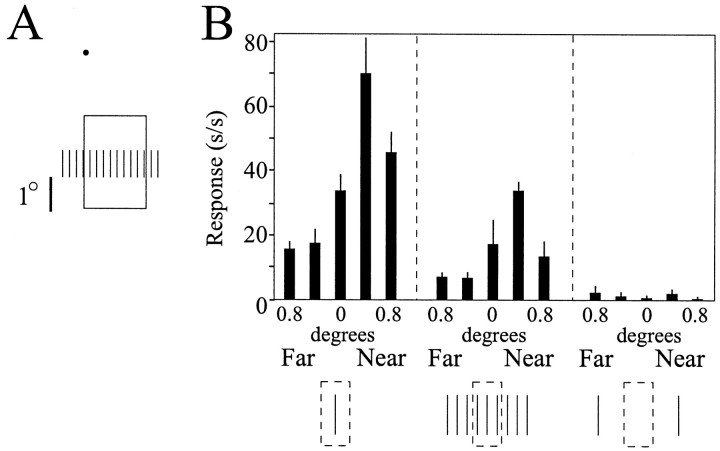
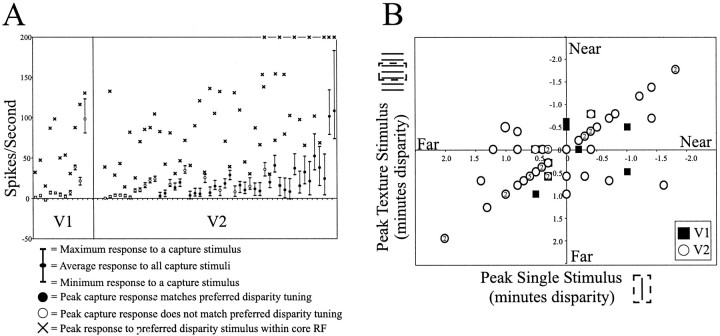
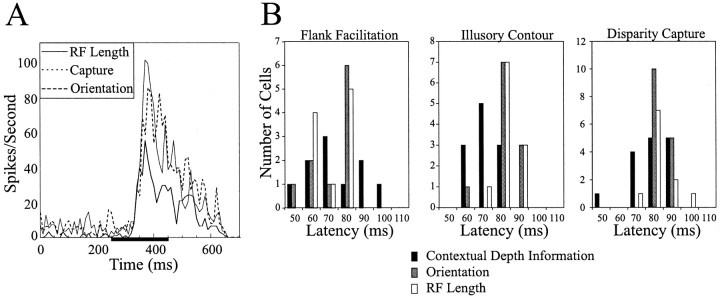
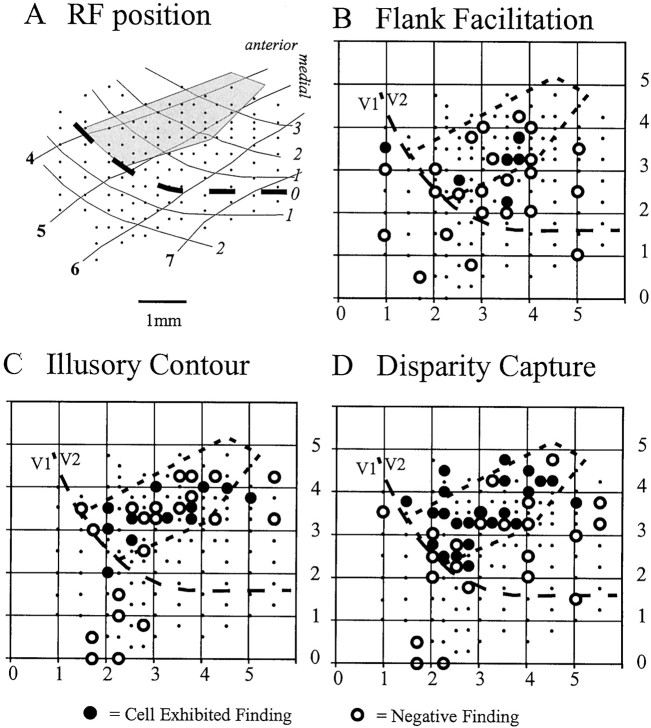
The visual system uses information about the relative depth of contours and surfaces to link and segment elements of visual scenes. The integration of form and depth information was studied in areas V1 and V2 of the alert macaque. Neurons in area V2 used contextual depth information to integrate occluded contours, signal the presence of object boundaries, and segment surfaces: (1) Amodal contour completion occurs when a contour passes behind an occluder. The basis of contour completion, the facilitation of neuronal responses to stimuli located within their receptive fields (RFs) by contextual lines lying outside their RFs, was blocked by orthogonal lines intersecting the contours but was recovered when the orthogonal line was placed in the near depth plane. (2) An illusory contour will modally complete separated elements located across an isoluminant field if the elements are placed in the near depth plane. V2 neurons responded when line segments were placed outside the RF in the near depth plane and a field of uniform luminance covered the RF. (3) Texture elements within a surface will "capture" the perceived depth consistent with the disparity of the surface's boundary, even when given no disparity themselves. V2 neurons responded to the center elements of a grating as if they contained disparity, even though disparity was present only for the grating's end elements located beyond the RF borders. These results, which were more common in V2 than in V1, demonstrate a role for V2 in the three-dimensional representation of surfaces in space.
Figures
References
-
- Allman JM, Miezin F, McGuiness E. Directions and velocity specific surround in three cortical visual areas of the owl monkey. Perception. 1985;14:105–126. - PubMed
-
- Bishop PO. Seeing with two eyes. Aust J Sci. 1970;32:383–391.
-
- Bishop PO. Neurophysiology of binocular single vision and stereopsis. In: Jung R, editor. Central processing of visual information. A: Integrative functions and comparative data. Springer; Heildelberg: 1973. pp. 255–305.
-
- Bishop PO. Stereopsis and fusion. Trans Ophthalmol Soc N Z. 1974;26:17–27. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources