

Molecular genetics of pattern formation in the inner ear: do compartment boundaries play a role?
- PMID: 11050198
- PMCID: PMC34338
- DOI: 10.1073/pnas.97.22.11700
Molecular genetics of pattern formation in the inner ear: do compartment boundaries play a role?
Abstract
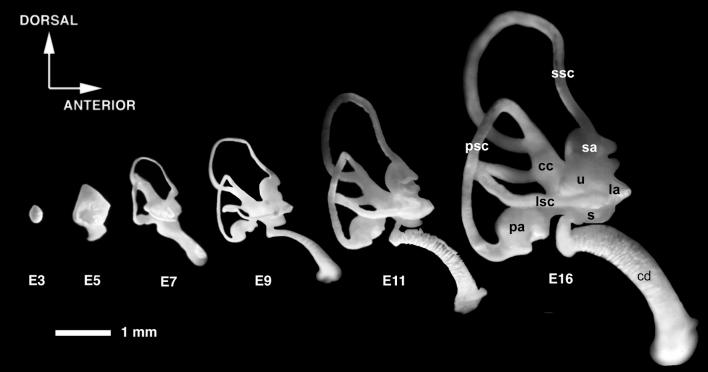
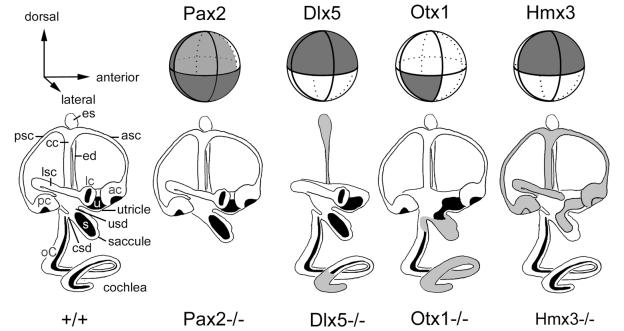
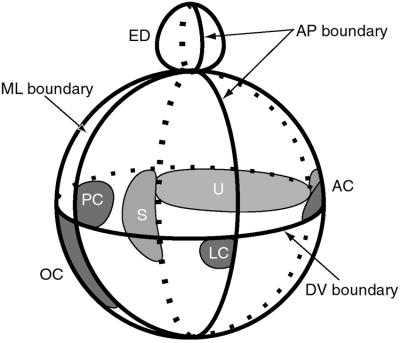
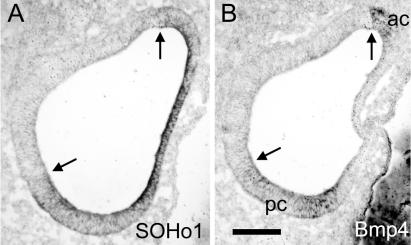
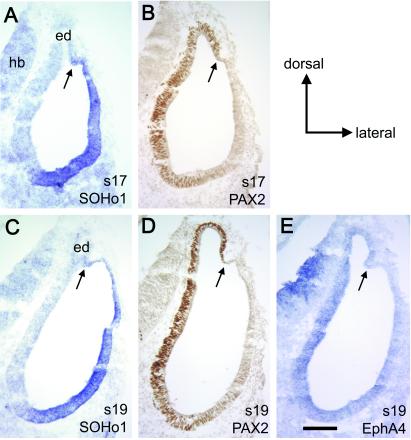
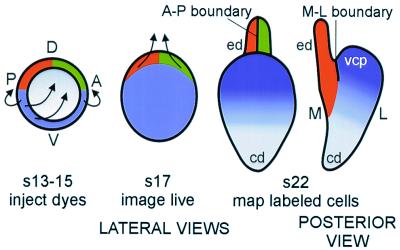
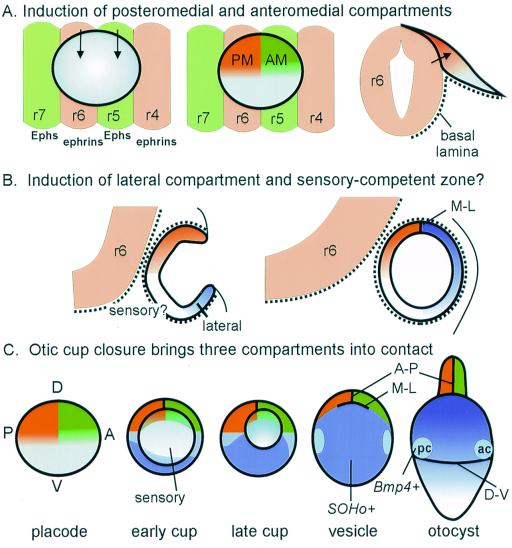
The membranous labyrinth of the inner ear establishes a precise geometrical topology so that it may subserve the functions of hearing and balance. How this geometry arises from a simple ectodermal placode is under active investigation. The placode invaginates to form the otic cup, which deepens before pinching off to form the otic vesicle. By the vesicle stage many genes expressed in the developing ear have assumed broad, asymmetrical expression domains. We have been exploring the possibility that these domains may reflect developmental compartments that are instrumental in specifying the location and identity of different parts of the ear. The boundaries between compartments are proposed to be the site of inductive interactions required for this specification. Our work has shown that sensory organs and the endolymphatic duct each arise near the boundaries of broader gene expression domains, lending support to this idea. A further prediction of the model, that the compartment boundaries will also represent lineage-restriction compartments, is supported in part by fate mapping the otic cup. Our data suggest that two lineage-restriction boundaries intersect at the dorsal pole of the otocyst, a convergence that may be critical for the specification of endolymphatic duct outgrowth. We speculate that the patterning information necessary to establish these two orthogonal boundaries may emanate, in part, from the hindbrain. The compartment boundary model of ear development now needs to be tested through a variety of experimental perturbations, such as the removal of boundaries, the generation of ectopic boundaries, and/or changes in compartment identity.
Figures
References
-
- Lang H, Bever M M, Fekete D M. J Comp Neurol. 2000;417:205–220. - PubMed
-
- Fekete D M. Curr Opin Neurobiol. 1996;6:533–541. - PubMed
-
- Whitfield T, Haddon C, Lewis J. Semin Cell Dev Biol. 1997;8:239–247. - PubMed
-
- Torres M, Giraldez F. Mech Dev. 1998;71:5–21. - PubMed
-
- Rivolta M N. Audiol Neurootol. 1997;2:36–49. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
