

Mechanical bases of frequency tuning and neural excitation at the base of the cochlea: comparison of basilar-membrane vibrations and auditory-nerve-fiber responses in chinchilla
- PMID: 11050204
- PMCID: PMC34344
- DOI: 10.1073/pnas.97.22.11744
Mechanical bases of frequency tuning and neural excitation at the base of the cochlea: comparison of basilar-membrane vibrations and auditory-nerve-fiber responses in chinchilla
Abstract
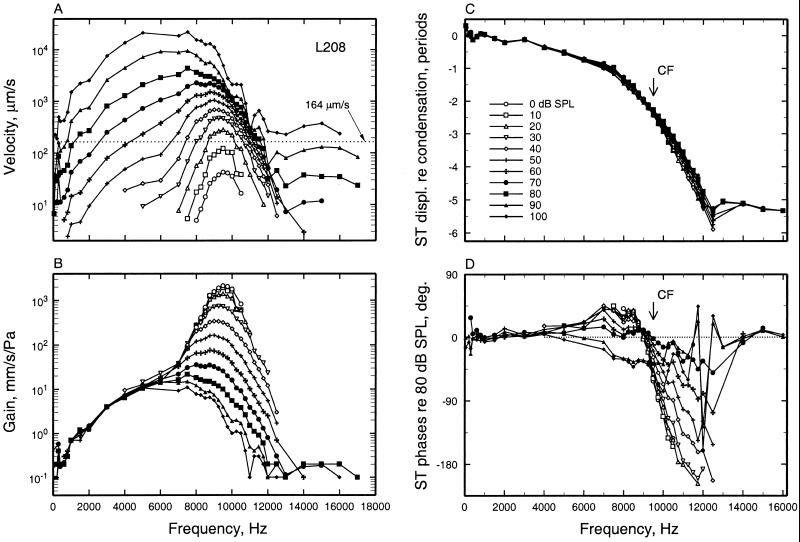
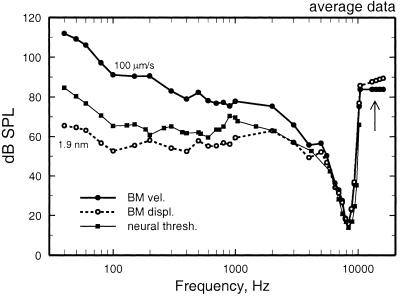
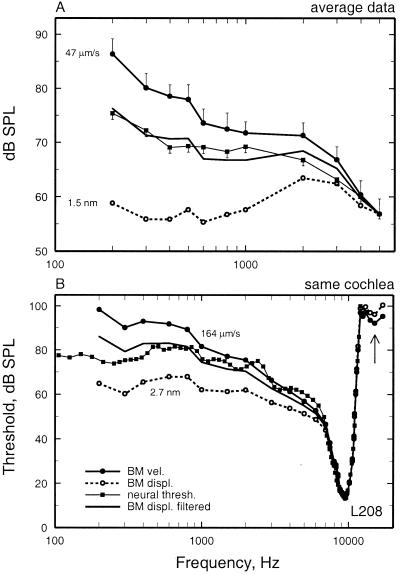
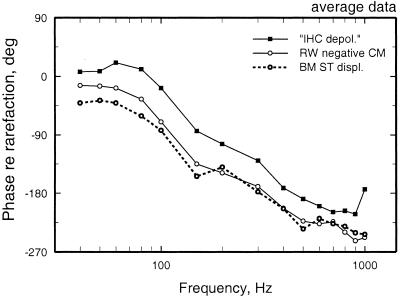
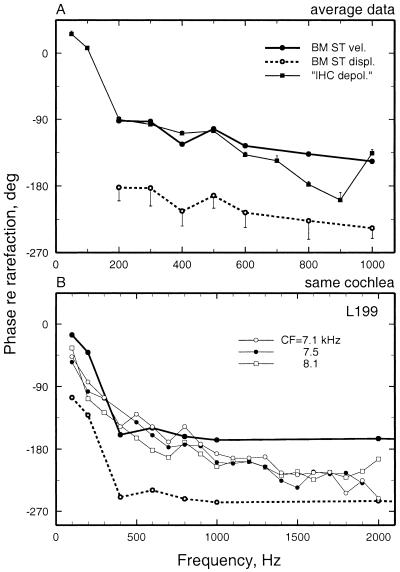
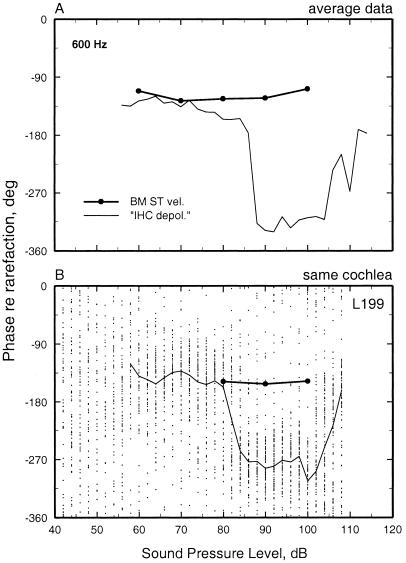
We review the mechanical origin of auditory-nerve excitation, focusing on comparisons of the magnitudes and phases of basilar-membrane (BM) vibrations and auditory-nerve fiber responses to tones at a basal site of the chinchilla cochlea with characteristic frequency approximately 9 kHz located 3.5 mm from the oval window. At this location, characteristic frequency thresholds of fibers with high spontaneous activity correspond to magnitudes of BM displacement or velocity in the order of 1 nm or 50 microm/s. Over a wide range of stimulus frequencies, neural thresholds are not determined solely by BM displacement but rather by a function of both displacement and velocity. Near-threshold, auditory-nerve responses to low-frequency tones are synchronous with peak BM velocity toward scala tympani but at 80-90 dB sound pressure level (in decibels relative to 20 microPascals) and at 100-110 dB sound pressure level responses undergo two large phase shifts approaching 180 degrees. These drastic phase changes have no counterparts in BM vibrations. Thus, although at threshold levels the encoding of BM vibrations into spike trains appears to involve only relatively minor signal transformations, the polarity of auditory-nerve responses does not conform with traditional views of how BM vibrations are transmitted to the inner hair cells. The response polarity at threshold levels, as well as the intensity-dependent phase changes, apparently reflect micromechanical interactions between the organ of Corti, the tectorial membrane and the subtectorial fluid, and/or electrical and synaptic processes at the inner hair cells.
Figures
References
-
- Spoendlin H. In: Physiology of the Ear. Jahn A F, Santos-Sacchi J, editors. New York: Raven; 1988. pp. 201–219.
-
- Russell I J, Richardson G P, Cody A R. Nature (London) 1986;321:517–519. - PubMed
-
- Ruggero M A. In: Active Hearing. Flock A, Ottoson D, Ulfendahl M, editors. Oxford: Pergamon; 1995. pp. 321–336.
-
- Sellick P M, Patuzzi R, Johnstone B M. J Acoust Soc Am. 1982;72:131–141. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
