

Molecular mechanisms of sound amplification in the mammalian cochlea
- PMID: 11050206
- PMCID: PMC34346
- DOI: 10.1073/pnas.97.22.11759
Molecular mechanisms of sound amplification in the mammalian cochlea
Abstract
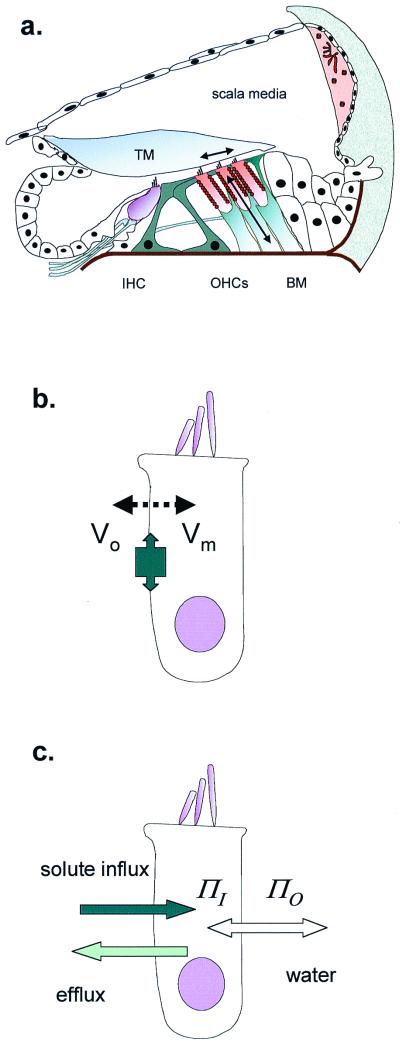
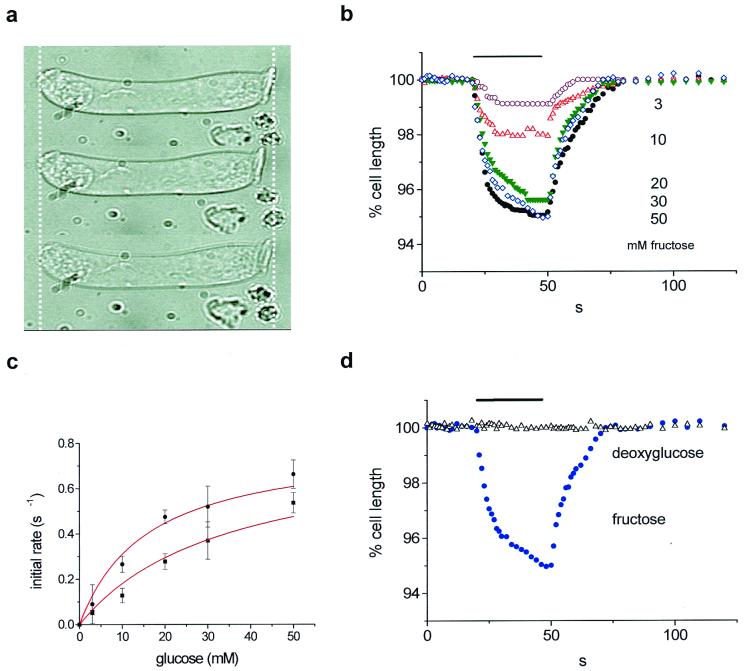
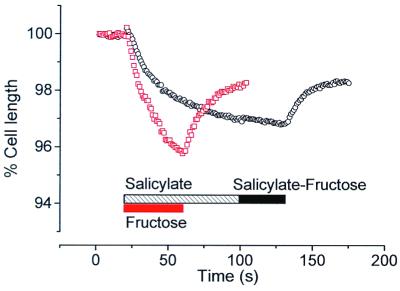
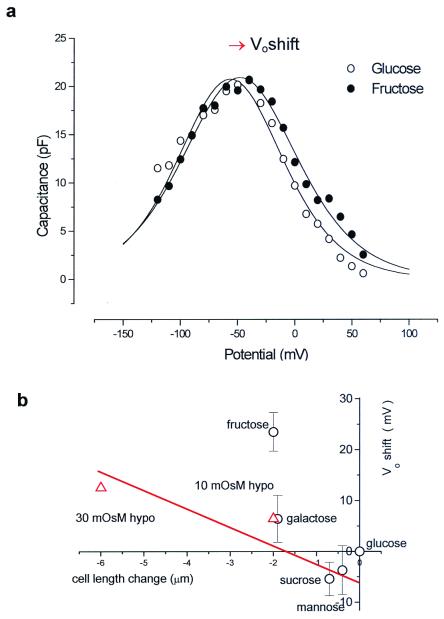
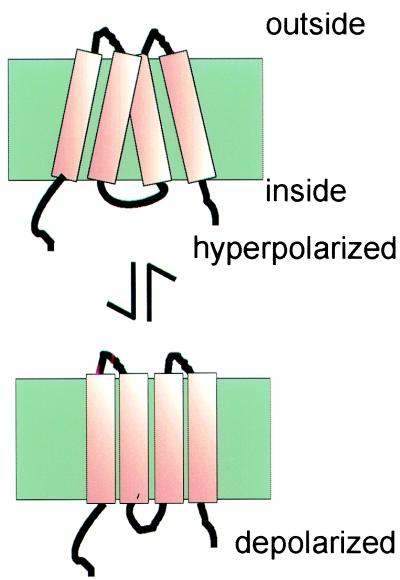
Mammalian hearing depends on the enhanced mechanical properties of the basilar membrane within the cochlear duct. The enhancement arises through the action of outer hair cells that act like force generators within the organ of Corti. Simple considerations show that underlying mechanism of somatic motility depends on local area changes within the lateral membrane of the cell. The molecular basis for this phenomenon is a dense array of particles that are inserted into the basolateral membrane and that are capable of sensing membrane potential field. We show here that outer hair cells selectively take up fructose, at rates high enough to suggest that a sugar transporter may be part of the motor complex. The relation of these findings to a recent candidate for the molecular motor is also discussed.
Figures
References
-
- Gold T. Proc R Soc London Ser B. 1948;135:492–498.
-
- Zweig G. J Acoust Soc Am. 1991;89:1229–1254. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
