

Detection of synchrony in the activity of auditory nerve fibers by octopus cells of the mammalian cochlear nucleus
- PMID: 11050208
- PMCID: PMC34348
- DOI: 10.1073/pnas.97.22.11773
Detection of synchrony in the activity of auditory nerve fibers by octopus cells of the mammalian cochlear nucleus
Abstract
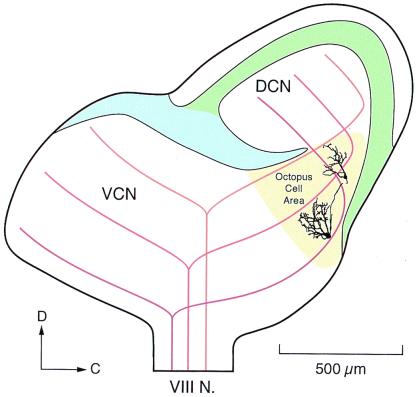
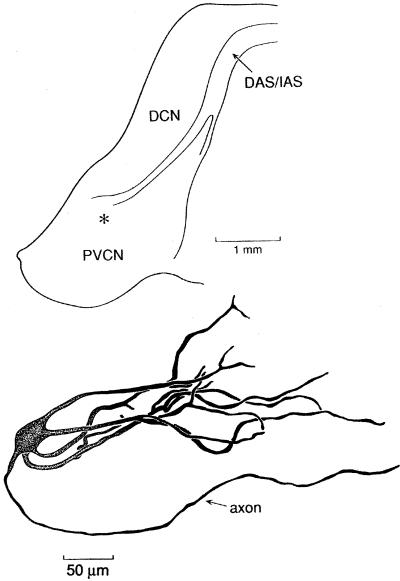
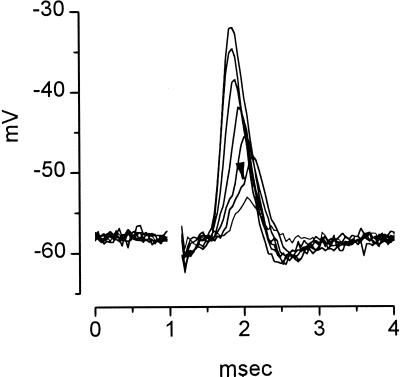
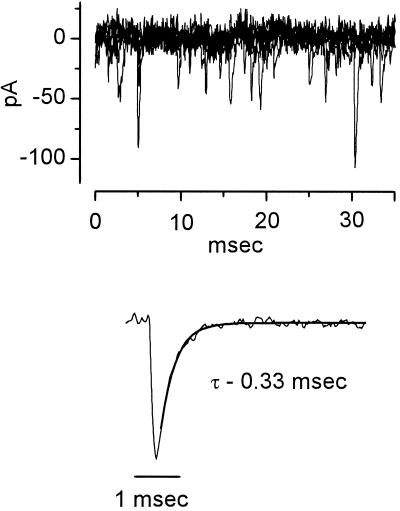
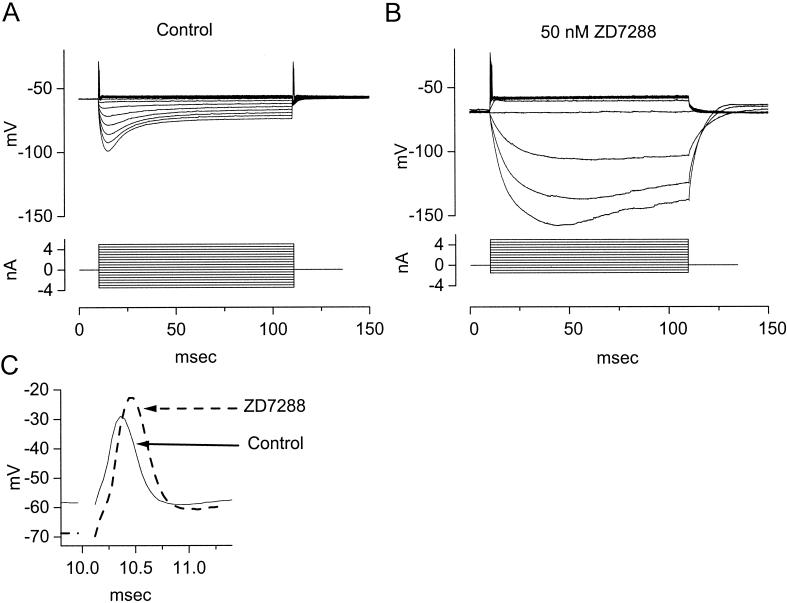
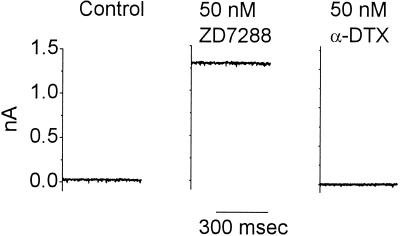
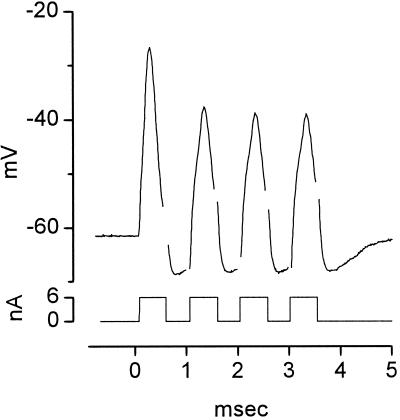
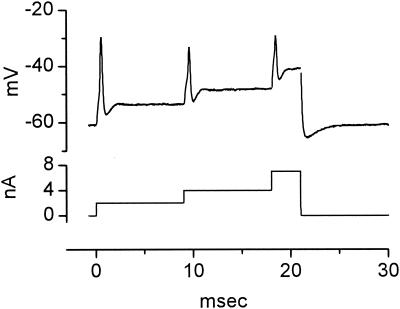
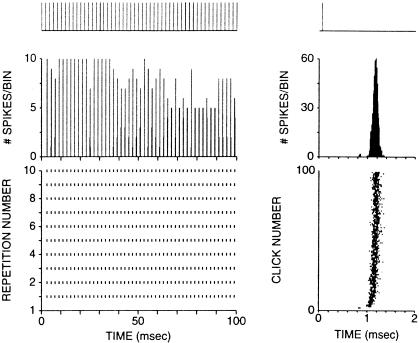
The anatomical and biophysical specializations of octopus cells allow them to detect the coincident firing of groups of auditory nerve fibers and to convey the precise timing of that coincidence to their targets. Octopus cells occupy a sharply defined region of the most caudal and dorsal part of the mammalian ventral cochlear nucleus. The dendrites of octopus cells cross the bundle of auditory nerve fibers just proximal to where the fibers leave the ventral and enter the dorsal cochlear nucleus, each octopus cell spanning about one-third of the tonotopic array. Octopus cells are excited by auditory nerve fibers through the activation of rapid, calcium-permeable, alpha-amino-3-hydroxy-5-methyl-4-isoxazole-propionate receptors. Synaptic responses are shaped by the unusual biophysical characteristics of octopus cells. Octopus cells have very low input resistances (about 7 M Omega), and short time constants (about 200 microsec) as a consequence of the activation at rest of a hyperpolarization-activated mixed-cation conductance and a low-threshold, depolarization-activated potassium conductance. The low input resistance causes rapid synaptic currents to generate rapid and small synaptic potentials. Summation of small synaptic potentials from many fibers is required to bring an octopus cell to threshold. Not only does the low input resistance make individual excitatory postsynaptic potentials brief so that they must be generated within 1 msec to sum but also the voltage-sensitive conductances of octopus cells prevent firing if the activation of auditory nerve inputs is not sufficiently synchronous and depolarization is not sufficiently rapid. In vivo in cats, octopus cells can fire rapidly and respond with exceptionally well-timed action potentials to periodic, broadband sounds such as clicks. Thus both the anatomical specializations and the biophysical specializations make octopus cells detectors of the coincident firing of their auditory nerve fiber inputs.
Figures
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
