

Review
doi: 10.1016/s0962-8924(00)01841-9.
Leukocytes navigate by compass: roles of PI3Kgamma and its lipid products
Affiliations
- PMID: 11050418
- PMCID: PMC2819116
- DOI: 10.1016/s0962-8924(00)01841-9
Item in Clipboard
Review
Leukocytes navigate by compass: roles of PI3Kgamma and its lipid products
Trends Cell Biol.
2000 Nov.
Abstract
Morphologic polarity is necessary for the motility of mammalian cells. In leukocytes responding to a chemoattractant, this polarity is regulated by activities of small Rho guanosine triphosphatases (Rho GTPases) and the phosphoinositide 3-kinases (PI3Ks). Moreover, in neutrophils, lipid products of PI3Ks appear to regulate activation of Rho GTPases, are required for cell motility and accumulate asymmetrically to the plasma membrane at the leading edge of polarized cells. By spatially regulating Rho GTPases and organizing the leading edge of the cell, PI3Ks and their lipid products could play pivotal roles not only in establishing leukocyte polarity but also as compass molecules that tell the cell where to crawl.
Figures
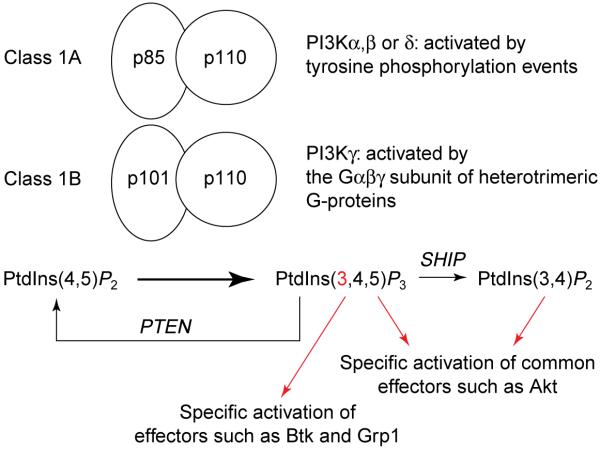
Class I phosphoinositide 3-kinases and the fate of their lipid product.
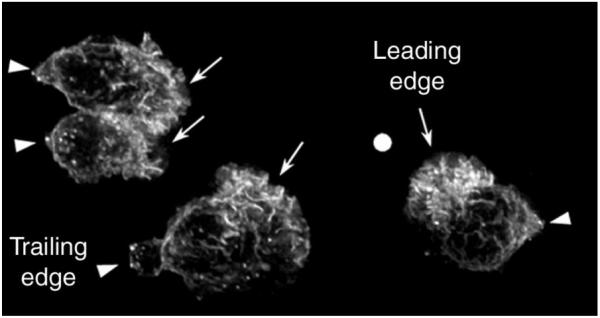
F-actin accumulation in chemoattractant-stimulated human neutrophils. When exposed to a chemotactic gradient, neutrophils polymerize F-actin asymmetrically to their pole facing the highest concentration of chemoattractant. In this case, human neutrophils were exposed to a gradient of f-Met-Leu-Phe (fMLP) delivered from a micropipette tip (shown by a white dot), allowed to polarize, fixed and their F-actin was revealed by fluorescence microscopy. These neutrophils exhibit a typical polarized morphology with a trailing edge (arrow heads) and an F-actin-enriched leading edge (arrows) facing up the gradient.
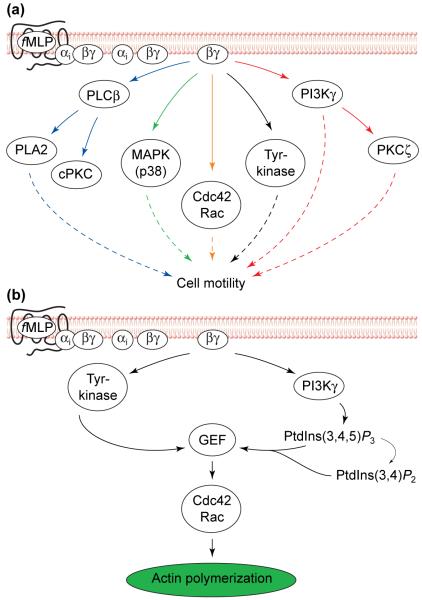
Overview of chemotactic signalling in neutrophils. (a) Several signalling pathways could regulate neutrophil motility. Chemoattractants, such as f-Met-Leu-Phe (fMLP), trigger signalling by activating their specific G-protein-coupled receptors (GPCRs) and Gi proteins at the surface of neutrophils, leading to release of the Gβγ subunit, which in turn activates directly or indirectly numerous signalling pathways in neutrophils, several of which have been suggested to regulate neutrophil motility (dotted arrows). (b) Putative signalling pathway regulating chemoattractant-induced activation of Rho GTPases in neutrophils. The Gβγ subunit, freed from the activated heterotrimeric G protein, activates phosphoinositide 3-kinase gamma (PI3Kγ) and cytosolic tyrosine kinases. The combined actions of tyrosine kinases and the PI3Kγ lipid products, phosphatidylinositol (3,4,5)-trisphosphate [PtdIns(3,4,5)P3] and/or phosphatidylinositol (3,4)-bisphosphate [PtdIns(3,4)P2], increase the activity of a specific GDP–GTP exchange factor (GEF; e.g. Vav) for the Rho GTPases. The activated GEF in turn activates the Rho GTPases Rac and Cdc42 that ultimately transmit signals to the actin-polymerization machinery. Abbreviations: cPKC, classical protein kinase C; MAPK, mitogen-activated protein kinase; PLA2, phospholipase A2; PLCβ, phospholipase C β.
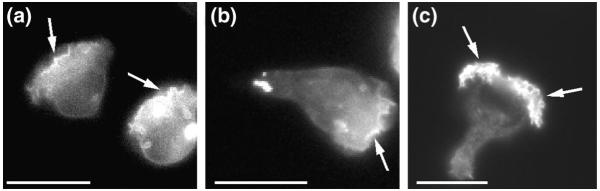
Localization of lipid products of PI3Ks in chemoattractant-stimulated HL-60 cells. (a) Cells expressing the pleckstrin-homology domain of the protein kinase AKT/PKB fused to green-fluorescent protein (PHAKT–GFP) migrate in a gradient of f-Met-Leu-Phe (fMLP) delivered from a micropipette tip (located outside of the field of view in the top left corner). Fluorescence microscopy shows PHAKT–GFP predominantly localized at the plasma membrane at the leading edge (arrows). Since PHAKT–GFP binds selectively to the phosphoinositide 3-kinase (PI3K) lipid products phosphatidylinositol (3,4,5)-trisphosphate [PtdIns(3,4,5)P3] and/or phosphatidylinositol (3,4)-bisphosphate [PtdIns(3,4)P2] at the plasma membrane, this indicates that one or both of these phospholipids accumulates at the leading edge of polarized neutrophils. (b) Even under conditions in which all the receptors are detecting the same ambient concentration of chemoattractant, after a uniform stimulation with fMLP, the cells polarize and show asymmetric accumulation of PHAKT–GFP at the leading edge (arrow). (c) PtdIns(3,4,5)P3 distribution on a chemoattractant-stimulated cell is revealed with the use of a specific antibody (from Echelon Research Laboratories, UT, USA) that does not bind to PtdIns(3,4)P2. In parallel to results obtained with PHAKT–GFP, this phospholipid is seen only at the leading edge of polarized cells (arrows). Bars, 15 μm.
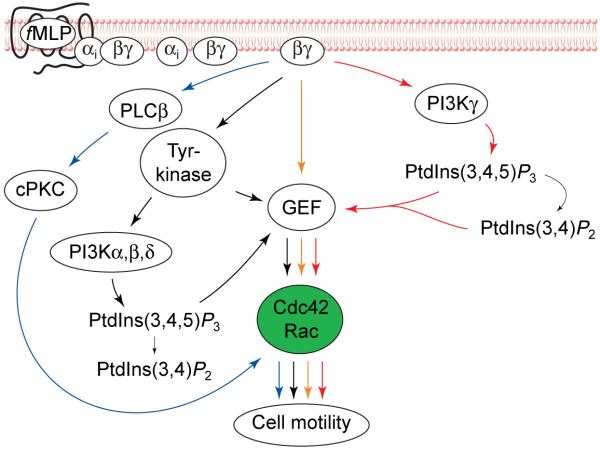
Alternative pathways for activation of Rho GTPases by chemoattractants in neutrophils. Pathways in addition to the phosphoinositide 3-kinase gamma (PI3Kγ) pathway (red arrows) could mediate activation of Rho GTPases in neutrophils. The activated cytosolic tyrosine kinases (black arrows) could induce activation of class 1A PI3Ks (α, β and δ; see Box 1). Combined effects of tyrosine kinases and PI3K lipid products on a GDP–GTP exchange factor (GEF) could induce Rac or Cdc42 activation. Alternatively, released Gβγ could also activate a GEF directly (yellow arrows). Gβγ also activates the phospholipases PLCβ2 and PLCβ3 in neutrophils (blue arrows). Activation of these phospholipases results in an increase in cytosolic Ca2+ and formation of diacylglycerol at the plasma membrane. These two events lead to the activation of classical protein kinase C (cPKC) enzymes. These protein kinases have been shown to activate Rac and Cdc42 independently of PI3K in neutrophils. Abbreviations: PtdIns(3,4,5)P3, phosphatidylinositol (3,4,5)-trisphosphate; PtdIns(3,4)P2, phosphatidylinositol (3,4)-bisphosphate.

Amplification of the accumulation of phosphoinositide 3-kinase (PI3K) lipid products relative to the gradient of chemoattractant determines cell polarity. (a) Amplification could occur in part through positive-feedback loops involving PI3Kγ and PI3Ks of class 1A. In the top scheme, chemoattractant-induced activation of cytosolic tyrosine kinases links activation of PI3Kγ to that of PI3Ks of class 1A. In this scenario, Gβγ activates PI3Kγ, which in turn produces phosphatidylinositol (3,4,5)-trisphosphate [PtdIns(3,4,5)P3]. This lipid product then somehow induces the activation of cytosolic tyrosine kinases that then relay the signal to PI3Ks of class 1A (α, β and δ). These PI3Ks then further increase the accumulation of PI3K lipid products. The increased production of PI3K lipid products and the effect of tyrosine kinases then increase the activity of a GDP–GTP exchange factor (GEF) and the Rho GTPases that induce cell polarity. In the bottom scheme, Rho GTPases link PI3Kγ to activation of class1A PI3Ks. Gβγ-induced activation of PI3Kγ and cytosolic tyrosine kinases lead to activation of the Rho GTPases, just as described in the legend of Fig. 2. These Rho GTPases not only induce cell polarity but also activate class 1A PI3Ks, which in turn further increase the level of PI3K lipid product formation. This increase in PI3K lipid products then leads to a greater increase in activity of Rho GTPases, which reinforces polarity. (b) Asymmetric accumulation of PI3K lipid products [including PtdIns(3,4,5)P3 and phosphatidylinositol (3,4)-bisphosphate (PtdIns(3,4)P2)] in polarized neutrophils also requires inhibition of signalling at the trailing edge. The positive-feedback loops increasing the level of formation of PI3K lipid products cannot explain their absence in the trailing edge of polarized cells. A model, suggests that activated receptors might also generate a long-range inhibitor (–) of signalling, in addition to local enhancers (+) such as the postulated PtdIns(3,4,5)P3-dependent positive feedback. The hypothetical inhibitor would initially prevent PI3K lipid product accumulation everywhere, but proportionally more at the pole of the cell facing the lowest concentration of chemoattractant in a gradient (I: initially). The combined effects of global inhibitors and local activators of PI3K lipid product formation would then result in accumulation of PI3K lipid products exclusively at the maximally stimulated area of the neutrophil plasma membrane. This local accumulation of PI3K lipid products would lead to cell polarization through a local activation of the Rho GTPases (II: after amplification).
References
-
- Wymann MP, Pirola L. Structure and function of phosphoinositide 3-kinases. Biochim. Biophys. Acta. 1998;1436:127–150. - PubMed
-
- Sasaki T, et al. Function of PI3Kγ in thymocyte development, T cell activation, and neutrophil migration. Science. 2000;287:1040–1046. - PubMed
-
- Li Z, et al. Roles of PLC-β2 and -β3 and PI3Kγ in chemoattractant-mediated signal transduction. Science. 2000;287:1046–1049. - PubMed
-
- Hirsch E, et al. Central role for G protein-coupled phosphoinositide 3-kinase γ in inflammation. Science. 2000;287:1049–1053. - PubMed
-
- Maier U, et al. Roles of non-catalytic subunits in Gβγ-induced activation of class I phosphoinositide 3-kinase isoforms β and γ. J. Biol. Chem. 1999;274:29311–29317. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
