

The effects of 1 alpha,25-dihydroxyvitamin D(3) on matrix metalloproteinase and prostaglandin E(2) production by cells of the rheumatoid lesion
- PMID: 11056661
- PMCID: PMC17774
- DOI: 10.1186/ar12
The effects of 1 alpha,25-dihydroxyvitamin D(3) on matrix metalloproteinase and prostaglandin E(2) production by cells of the rheumatoid lesion
Abstract
Introduction: 1alpha,25-dihydroxyvitamin D(3)[1alpha,25(OH)(2)D(3)], the biologically active metabolite of vitamin D3, acts through an intracellular vitamin D receptor (VDR) and has several immunostimulatory effects. Animal studies have shown that production of some matrix metalloproteinases (MMPs) may be upregulated in rat chondrocytes by administration of 1alpha,25(OH)(2)D(3); and cell cultures have suggested that 1alpha,25(OH)(2)D(3) may affect chondrocytic function. Discoordinate regulation by vitamin D of MMP-1 and MMP-9 in human mononuclear phagocytes has also been reported. These data suggest that vitamin D may regulate MMP expression in tissues where VDRs are expressed. Production of 1alpha,25(OH)(2)D(3) within synovial fluids of arthritic joints has been shown and VDRs have been found in rheumatoid synovial tissues and at sites of cartilage erosion. The physiological function of 1alpha,25(OH)(2)D(3) at these sites remains obscure. MMPs play a major role in cartilage breakdown in the rheumatoid joint and are produced locally by several cell types under strict control by regulatory factors. As 1alpha,25(OH)(2)D(3) modulates the production of specific MMPs and is produced within the rheumatoid joint, the present study investigates its effects on MMP and prostaglandin E(2) (PGE(2)) production in two cell types known to express chondrolytic enzymes.
Aims: To investigate VDR expression in rheumatoid tissues and to examine the effects of 1alpha,25-dihydroxyvitamin D(3) on cultured rheumatoid synovial fibroblasts (RSFs) and human articular chondrocytes (HACs) with respect to MMP and PGE(2) production.
Methods: Rheumatoid synovial tissues were obtained from arthroplasty procedures on patients with late-stage rheumatoid arthritis; normal articular cartilage was obtained from lower limb amputations. Samples were embedded in paraffin, and examined for presence of VDRs by immunolocalisation using a biotinylated antibody and alkaline-phosphatase-conjugated avidin-biotin complex system. Cultured synovial fibroblasts and chrondrocytes were treated with either 1alpha,25(OH)(2)D(3) or interleukin (IL)-1beta, or both. Conditioned medium was assayed for MMP and PGE(2) by enzyme-linked immunosorbent assay (ELISA), and the results were normalised relative to control values.
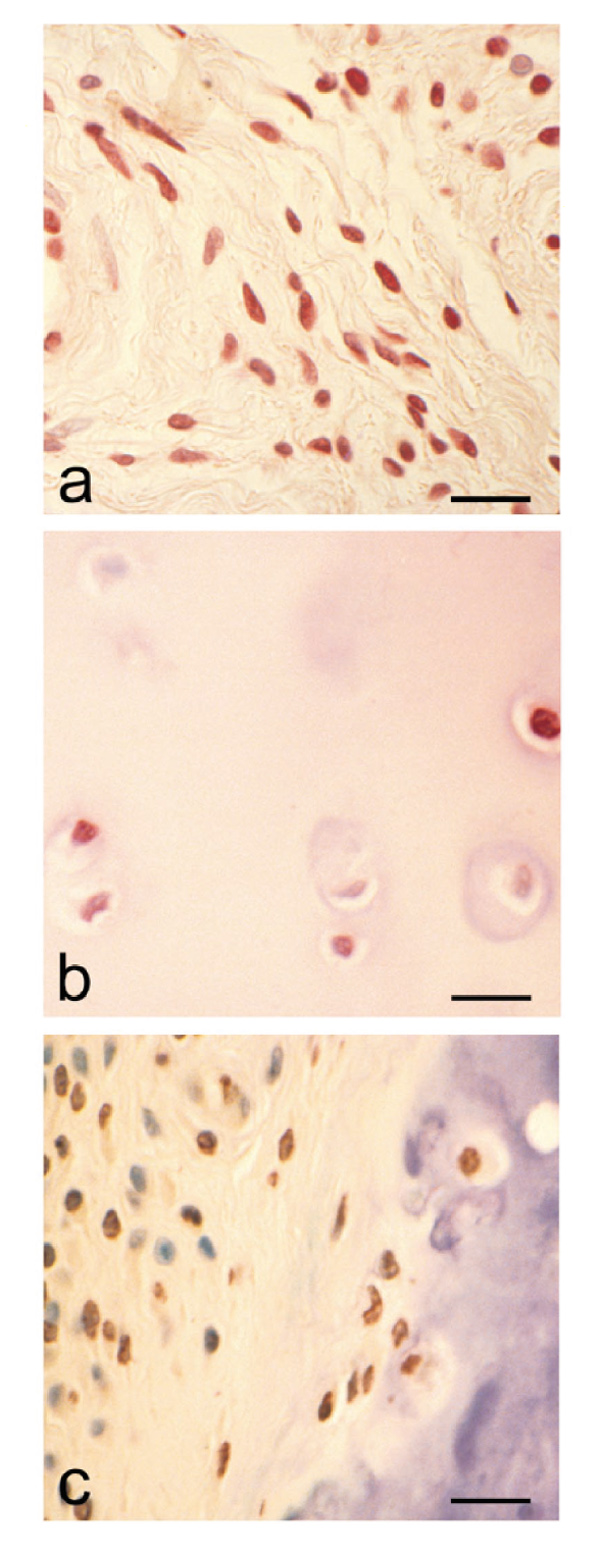
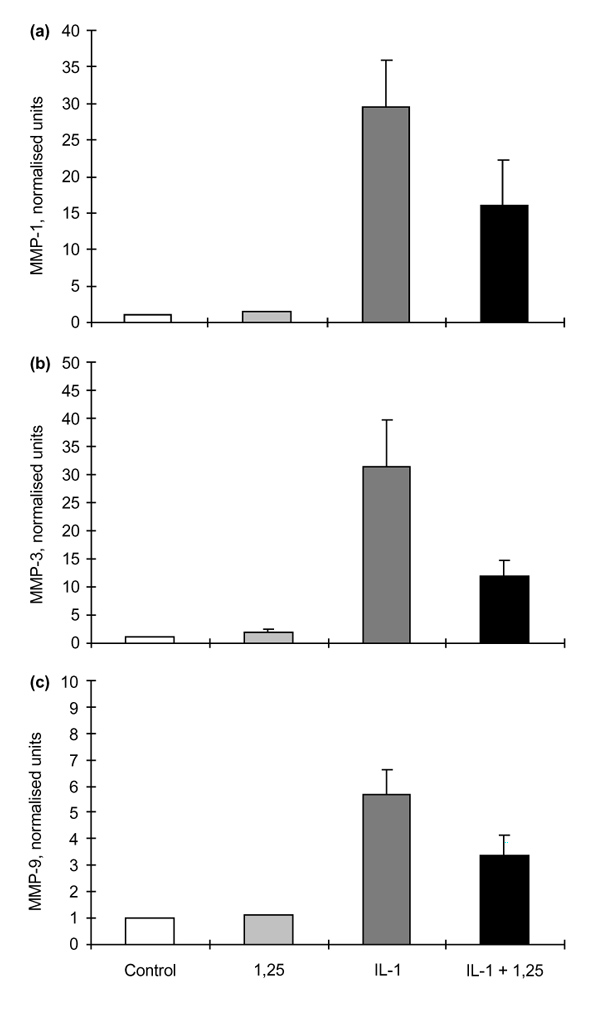
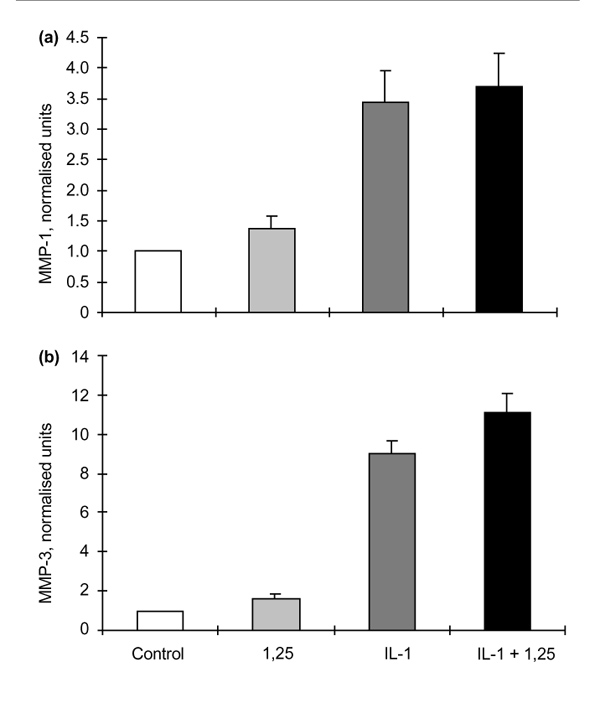
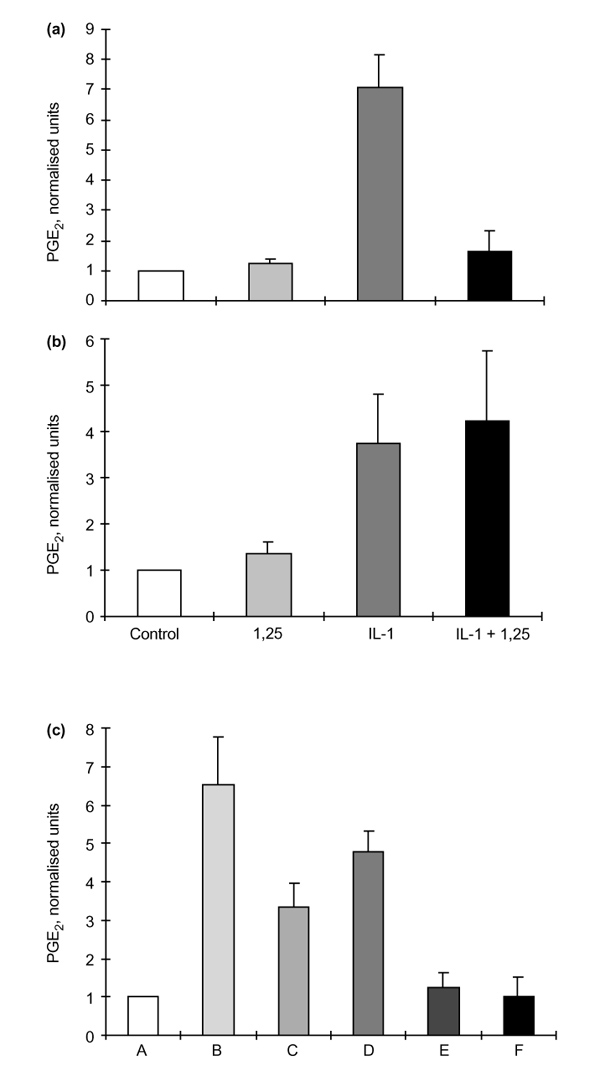
Results: The rheumatoid synovial tissue specimens (n=18) immunostained for VDRs showed positive staining but at variable distributions and in no observable pattern. VDR-positive cells were also observed in association with some cartilage-pannus junctions (the rheumatoid lesion). MMP production by RSFs in monolayer culture was not affected by treatment with 1alpha,25(OH)(2)D(3) alone, but when added simultaneously with IL-1beta the stimulation by IL-1beta was reduced from expected levels by up to 50%. In contrast, 1alpha,25(OH)(2)D(3) had a slight stimulatory effect on basal production of MMPs 1 and 3 by monolayer cultures of HACs, but stimulation of MMP-1 by IL-1beta was not affected by the simultaneous addition of 1alpha,25(OH)(2)D(3) whilst MMP-3 production was enhanced (Table 1). The production of PGE(2) by RSFs was unaffected by 1alpha,25(OH)(2)D(3) addition, but when added concomitantly with IL-1beta the expected IL-1beta-stimulated increase was reduced to almost basal levels. In contrast, IL-1beta stimulation of PGE(2) in HACs was not affected by the simultaneous addition of 1alpha,25(OH)(2)D(3)(Table 2). Pretreatment of RSFs with 1alpha,25(OH)(2)D(3) for 1h made no significant difference to IL-1beta-induced stimulation of PGE(2), but incubated for 16h suppressed the expected increase in PGE(2) to control values. This effect was also noted when 1alpha,25(OH)(2)D(3) was removed after the 16h and the IL-1beta added alone. Thus it appears that 1alpha,25(OH)(2)D(3) does not interfere with the IL-1beta receptor, but reduces the capacity of RSFs to elaborate PGE(2) after IL-1beta induction. Cells within the rheumatoid lesion which expressed VDR were fibroblasts, macrophages, lymphocytes and endothelial cells. These cells are thought to be involved in the degradative processes associated with rheumatoid arthritis (RA), thus providing evidence of a functional role of 1apha,25(OH)(2)D(3) in RA. MMPs may play important roles in the chondrolytic processes of the rheumatoid lesion and are known to be produced by both fibroblasts and chondrocytes. The 1alpha,25(OH)(2)D(3) had little effect of basal MMP production by RSFs, although more pronounced differences were noted when IL-1beta-stimulated cells were treated with 1alpha,25(OH)(2)D(3), with the RSF and HAC showing quite disparate responses. These opposite effects may be relevant to the processes of joint destruction, especially cartilage loss, as the ability of 1alpha,25(OH)(2)D(3) to potentiate MMP-1 and MMP-3 expression by 'activated' chondrocytes might facilitate intrinsic cartilage chondrolysis in vivo. By contrast, the MMP-suppressive effects observed for 1alpha,25(OH)2D3 treatment of 'activated' synovial fibroblasts might reduce extrinsic chondrolysis and also matrix degradation within the synovial tissue. Prostaglandins have a role in the immune response and inflammatory processes associated with RA. The 1alpha,25(OH)2D3 had little effect on basal PGE2 production by RSF, but the enhanced PGE2 production observed following IL-1beta stimulation of these cells was markedly suppressed by the concomitant addition of 1alpha,25(OH)2D3. As with MMP production, there are disparate effects of 1alpha,25(OH)2D3 on IL-1beta stimulated PGE2 production by the two cell types; 1alpha,25(OH)2D3 added concomitantly with IL-1beta had no effect on PGE2 production by HACs. In summary, the presence of VDRs in the rheumatoid lesion demonstrates that 1alpha,25(OH)2D3 may have a functional role in the joint disease process. 1 alpha,25(OH)2D3 does not appear to directly affect MMP or PGE2 production but does modulate cytokine-induced production.
Figures
Similar articles
-
Mast cell activation and its relation to proinflammatory cytokine production in the rheumatoid lesion.Arthritis Res. 2000;2(1):65-74. doi: 10.1186/ar70. Arthritis Res. 2000. PMID: 11219391 Free PMC article.
-
Expression of vitamin D receptors and matrix metalloproteinases in osteoarthritic cartilage and human articular chondrocytes in vitro.Osteoarthritis Cartilage. 2001 Jul;9(5):423-31. doi: 10.1053/joca.2000.0408. Osteoarthritis Cartilage. 2001. PMID: 11467890
-
Effect of histamine on the production of matrix metalloproteinases-1, -3, -8 and -13, and TNFalpha and PGE(2) by human articular chondrocytes and synovial fibroblasts in vitro: a comparative study.Virchows Arch. 2004 Nov;445(5):485-90. doi: 10.1007/s00428-004-1109-y. Epub 2004 Sep 17. Virchows Arch. 2004. PMID: 15378360
-
The role of cytokines in osteoarthritis pathophysiology.Biorheology. 2002;39(1-2):237-46. Biorheology. 2002. PMID: 12082286 Review.
-
Vitamin D, Autoimmune Disease and Rheumatoid Arthritis.Calcif Tissue Int. 2020 Jan;106(1):58-75. doi: 10.1007/s00223-019-00577-2. Epub 2019 Jul 8. Calcif Tissue Int. 2020. PMID: 31286174 Free PMC article. Review.
Cited by
-
Why do so many trials of vitamin D supplementation fail?Endocr Connect. 2020 Oct;9(9):R195-R206. doi: 10.1530/EC-20-0274. Endocr Connect. 2020. PMID: 33052876 Free PMC article. Review.
-
Effects of Vitamin D Supplementation during the Induction and Progression of Osteoarthritis in a Rat Model.Evid Based Complement Alternat Med. 2012;2012:156563. doi: 10.1155/2012/156563. Epub 2012 Oct 14. Evid Based Complement Alternat Med. 2012. PMID: 23118784 Free PMC article.
-
Calcium and vitamin D supplementation and incident rheumatoid arthritis: the Women's Health Initiative Calcium plus Vitamin D trial.Rheumatol Int. 2012 Dec;32(12):3823-30. doi: 10.1007/s00296-011-2268-1. Epub 2011 Dec 22. Rheumatol Int. 2012. PMID: 22190273 Free PMC article. Clinical Trial.
-
1,25(OH)2D3 and dexamethasone additively suppress synovial fibroblast activation by CCR6+ T helper memory cells and enhance the effect of tumor necrosis factor alpha blockade.Arthritis Res Ther. 2018 Sep 20;20(1):212. doi: 10.1186/s13075-018-1706-9. Arthritis Res Ther. 2018. PMID: 30236152 Free PMC article.
-
Calcipotriol counteracts betamethasone-induced decrease in extracellular matrix components related to skin atrophy.Arch Dermatol Res. 2014 Oct;306(8):719-29. doi: 10.1007/s00403-014-1485-3. Epub 2014 Jul 16. Arch Dermatol Res. 2014. PMID: 25027750 Free PMC article.
References
-
- Norman AW, Roth J, Orci L. The vitamin D endocrine system: steroid metabolism, hormone receptors and biological response (calcium binding proteins). Endocr Rev. 1982;3:331–336. - PubMed
-
- Suda T. The role of 1α25dihydroxyvitamin D3 in myeloid cell differentiation. Proc Soc Exp Biol Med. 1989;191:214–220. - PubMed
-
- Lemire JM. Immunomodulatory actions of 1,25dihydroxyvitamin D3. J Ster Biochem Mol Biol. 1995;53:599–602. - PubMed
-
- Lemire JM. Immunomodulatory role of 1,25dihydroxyvitmain D3. J Cell Biochem. 1992;49:26–31. - PubMed
-
- Gerstenfeld LC, Kelly CM, von Deck M, Lian JB. Effect of 1,25dihydroxyvitamin D3 on induction of chondrocyte maturation in culture: extracellular matrix gene expression and morphology. Endocrinology. 1990;126:1599–1609. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
