

The endoplasmic reticulum as one continuous Ca(2+) pool: visualization of rapid Ca(2+) movements and equilibration
- PMID: 11060024
- PMCID: PMC305795
- DOI: 10.1093/emboj/19.21.5729
The endoplasmic reticulum as one continuous Ca(2+) pool: visualization of rapid Ca(2+) movements and equilibration
Abstract
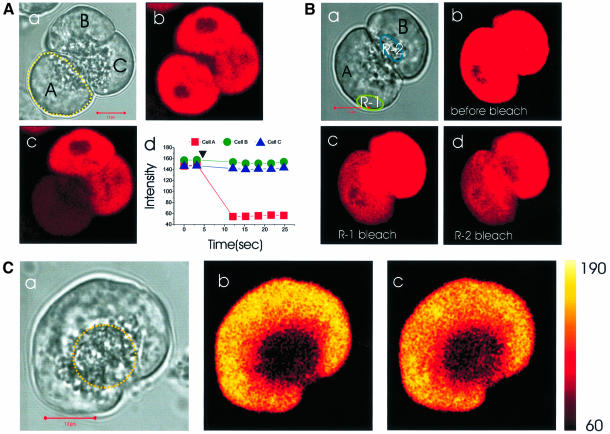
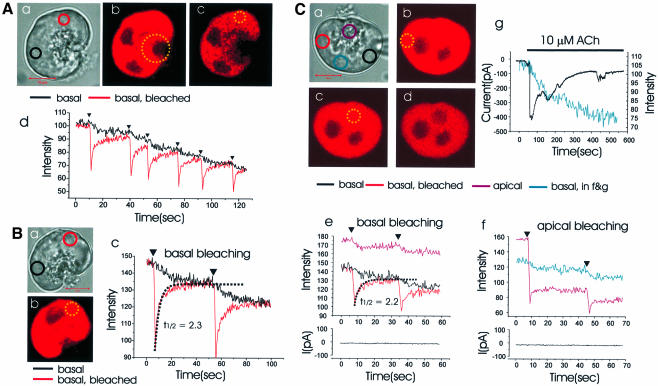
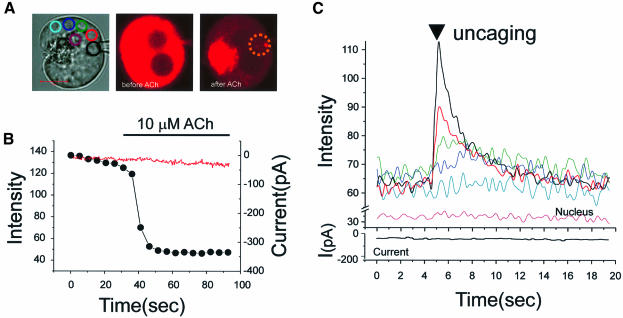
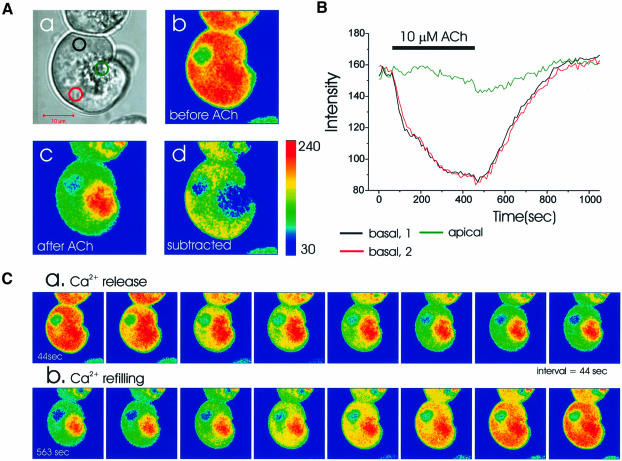
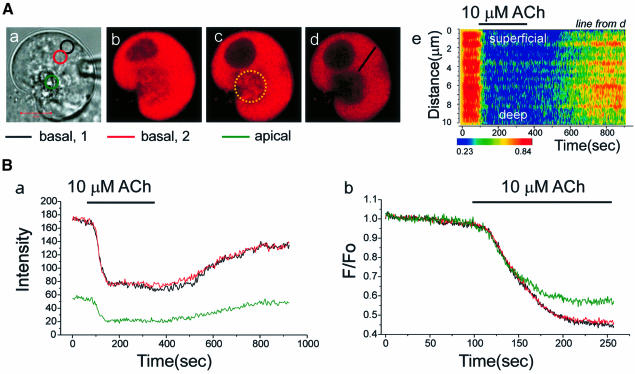
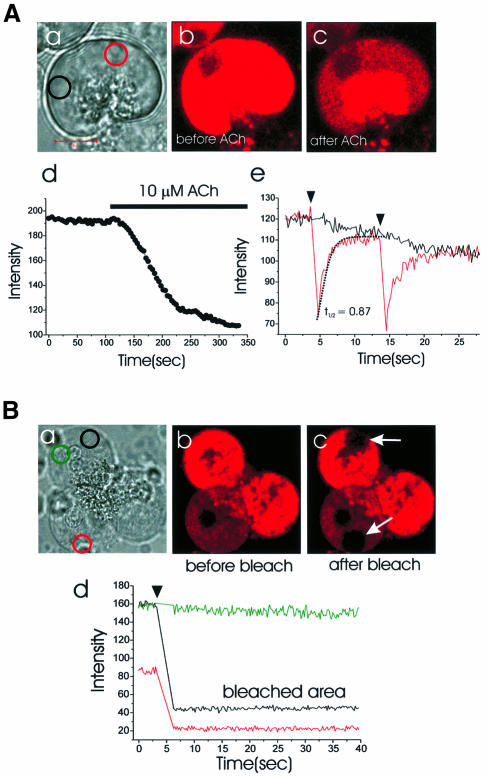
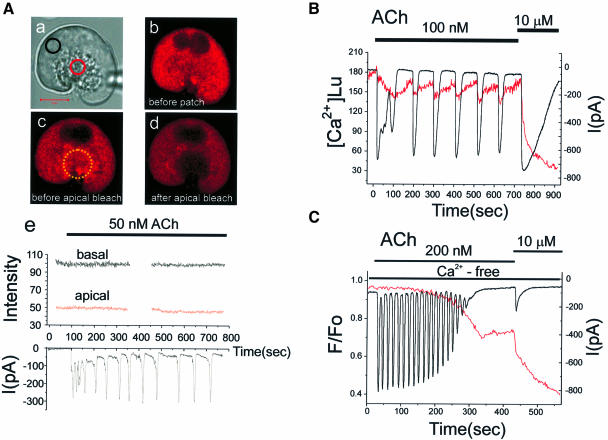
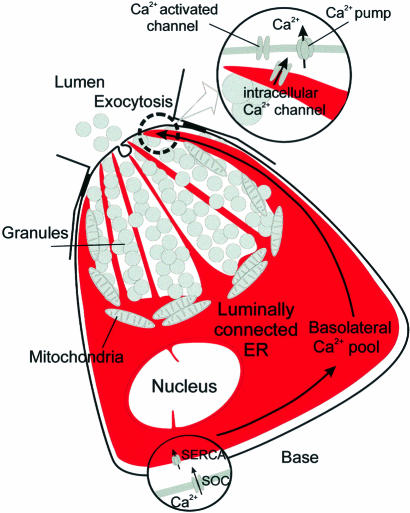
We investigated whether the endoplasmic reticulum (ER) is a functionally connected Ca(2+) store or is composed of separate subunits by monitoring movements of Ca(2+) and small fluorescent probes in the ER lumen of pancreatic acinar cells, using confocal microscopy, local bleaching and uncaging. We observed rapid movements and equilibration of Ca(2+) and the probes. The bulk of the ER at the base was not connected to the granules in the apical part, but diffusion into small apical ER extensions occurred. The connectivity of the ER Ca(2+) store was robust, since even supramaximal acetylcholine (ACh) stimulation for 30 min did not result in functional fragmentation. ACh could elicit a uniform decrease in the ER Ca(2+) concentration throughout the cell, but repetitive cytosolic Ca(2+) spikes, induced by a low ACh concentration, hardly reduced the ER Ca(2+) level. We conclude that the ER is a functionally continuous unit, which enables efficient Ca(2+) liberation. Ca(2+) released from the apical ER terminals is quickly replenished from the bulk of the rough ER at the base.
Figures
Similar articles
-
Store-operated Ca2+ entry in astrocytes: different spatial arrangement of endoplasmic reticulum explains functional diversity in vitro and in situ.Cell Calcium. 2008 Jun;43(6):591-601. doi: 10.1016/j.ceca.2007.10.004. Epub 2007 Dec 3. Cell Calcium. 2008. PMID: 18054077
-
Effects of elevated cytoplasmic calcium and protein kinase C on endoplasmic reticulum structure and function in HEK293 cells.Cell Calcium. 2000 Mar;27(3):175-85. doi: 10.1054/ceca.2000.0108. Cell Calcium. 2000. PMID: 11007130
-
The structural integrity of the endoplasmic reticulum, and its possible regulation by inositol 1,3,4,5-tetrakisphosphate.Cell Calcium. 2005 Aug;38(2):153-9. doi: 10.1016/j.ceca.2005.05.002. Cell Calcium. 2005. PMID: 16023721
-
Polarity in intracellular calcium signaling.Bioessays. 1999 Oct;21(10):851-60. doi: 10.1002/(SICI)1521-1878(199910)21:10<851::AID-BIES7>3.0.CO;2-F. Bioessays. 1999. PMID: 10497335 Review.
-
Ca²⁺ signalling in the endoplasmic reticulum/secretory granule microdomain.Cell Calcium. 2015 Oct;58(4):397-404. doi: 10.1016/j.ceca.2015.01.006. Epub 2015 Jan 25. Cell Calcium. 2015. PMID: 25662795 Review.
Cited by
-
Calcium transfer from the ER to other organelles for optimal signaling in Toxoplasma gondii.bioRxiv [Preprint]. 2024 Dec 5:2024.08.15.608087. doi: 10.1101/2024.08.15.608087. bioRxiv. 2024. PMID: 39185237 Free PMC article. Preprint.
-
Perinuclear, perigranular and sub-plasmalemmal mitochondria have distinct functions in the regulation of cellular calcium transport.EMBO J. 2001 Apr 17;20(8):1863-74. doi: 10.1093/emboj/20.8.1863. EMBO J. 2001. PMID: 11296220 Free PMC article.
-
Calcium regulation of EGF-induced ERK5 activation: role of Lad1-MEKK2 interaction.PLoS One. 2010 Sep 7;5(9):e12627. doi: 10.1371/journal.pone.0012627. PLoS One. 2010. PMID: 20830310 Free PMC article.
-
The Ca2+ concentration of the endoplasmic reticulum is a key determinant of ceramide-induced apoptosis: significance for the molecular mechanism of Bcl-2 action.EMBO J. 2001 Jun 1;20(11):2690-701. doi: 10.1093/emboj/20.11.2690. EMBO J. 2001. PMID: 11387204 Free PMC article.
-
IP(3) receptor subtype-dependent activation of store-operated calcium entry through I(CRAC).Cell Calcium. 2009 Apr;45(4):326-30. doi: 10.1016/j.ceca.2008.12.001. Epub 2009 Jan 20. Cell Calcium. 2009. PMID: 19157540 Free PMC article.
References
-
- Belan P.V., Gerasimenko,O.V., Tepikin,A.V. and Petersen,O.H. (1996) Localization of Ca2+ extrusion sites in pancreatic acinar cells. J. Biol. Chem., 271, 7615–7619. - PubMed
-
- Berridge M.J. (1993) Inositol trisphosphate and calcium signalling. Nature, 361, 315–325. - PubMed
-
- Berridge M.J. and Irvine,R.F. (1989) Inositol phosphates and cell signalling. Nature, 341, 197–205. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
