

Progression of a loop-loop complex to a four-way junction is crucial for the activity of a regulatory antisense RNA
- PMID: 11060041
- PMCID: PMC305787
- DOI: 10.1093/emboj/19.21.5905
Progression of a loop-loop complex to a four-way junction is crucial for the activity of a regulatory antisense RNA
Abstract
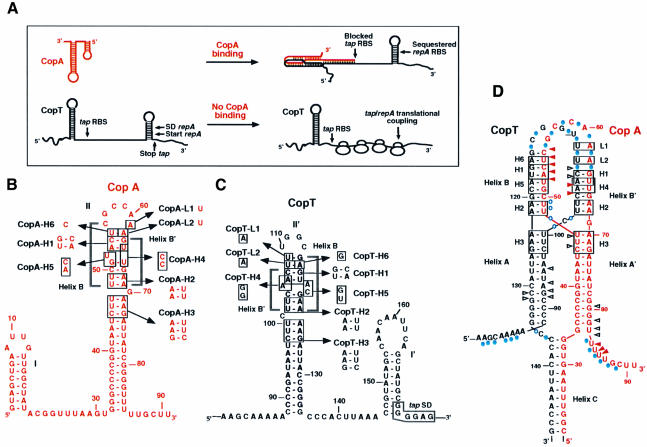
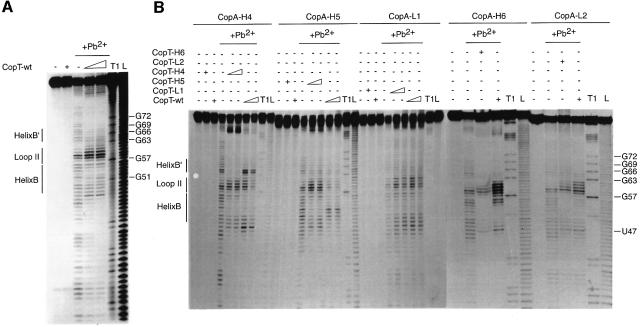
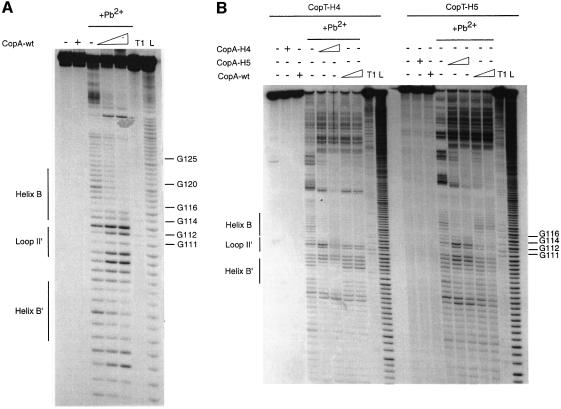
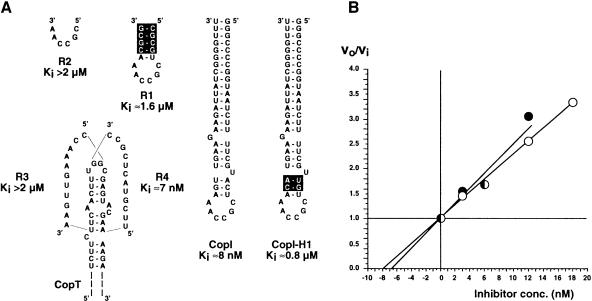
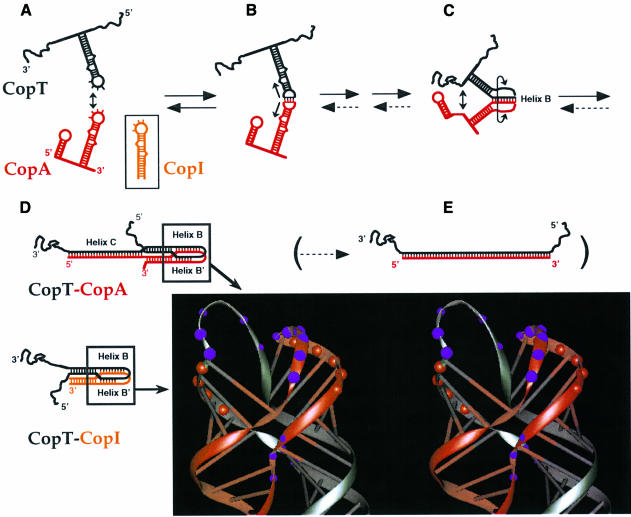
The antisense RNA, CopA, regulates the replication frequency of plasmid R1 through inhibition of RepA translation by rapid and specific binding to its target RNA (CopT). The stable CopA-CopT complex is characterized by a four-way junction structure and a side-by-side alignment of two long intramolecular helices. The significance of this structure for binding in vitro and control in vivo was tested by mutations in both CopA and CopT. High rates of stable complex formation in vitro and efficient inhibition in vivo required initial loop-loop complexes to be rapidly converted to extended interactions. These interactions involve asymmetric helix progression and melting of the upper stems of both RNAs to promote the formation of two intermolecular helices. Data presented here delineate the boundaries of these helices and emphasize the need for unimpeded helix propagation. This process is directional, i.e. one of the two intermolecular helices (B) must form first to allow formation of the other (B'). A binding pathway, characterized by a hierarchy of intermediates leading to an irreversible and inhibitory RNA-RNA complex, is proposed.
Figures
References
-
- Asano K. and Mizobuchi,K. (2000) Structural analysis of late intermediate complex formed between plasmid ColIb-P9 Inc RNA and its target RNA. J. Biol. Chem., 275, 1269–1274. - PubMed
-
- Asano K., Kato,A., Morikawi,H., Hama,C., Shiba,K. and Mizobuchi,K. (1991) Positive and negative regulations of plasmid ColIb-P9 repZ gene expression at the translational level. J. Biol. Chem., 266, 3774–3781. - PubMed
-
- Asano K., Niimi,T., Yokoyama,S. and Mizobuchi,K. (1998) Structural basis for binding of the plasmid ColIb-P9 antisense Inc RNA to its target RNA with the 5′-rUUGGCG-3′ motif in the loop sequence. J. Biol. Chem., 273, 11826–11838. - PubMed
-
- Berzal-Herranz A., Wagner,E.G.H. and Diaz-Orejas,R. (1991) Control of replication of plasmid R1: the intergenic region between copA and repA modulates the level of expression of repA. Mol. Microbiol., 5, 97–108. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
