

Polypeptide release by Hsp90 involves ATP hydrolysis and is enhanced by the co-chaperone p23
- PMID: 11060043
- PMCID: PMC305790
- DOI: 10.1093/emboj/19.21.5930
Polypeptide release by Hsp90 involves ATP hydrolysis and is enhanced by the co-chaperone p23
Abstract
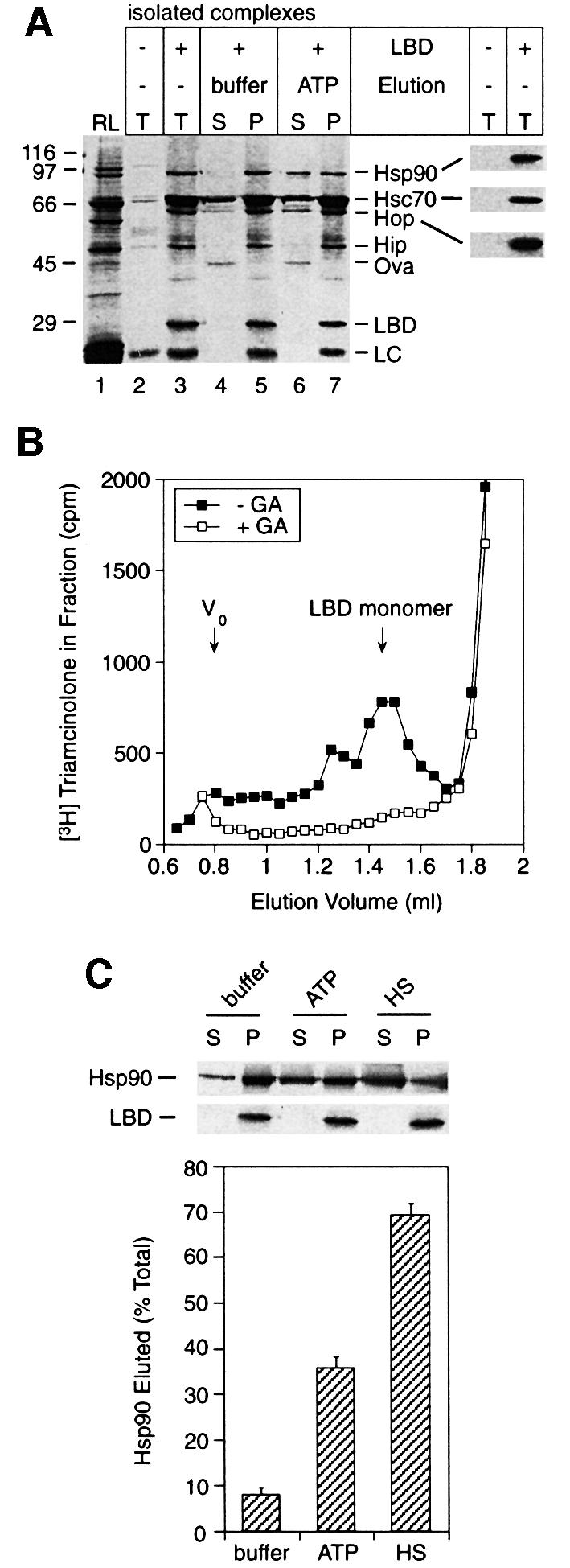
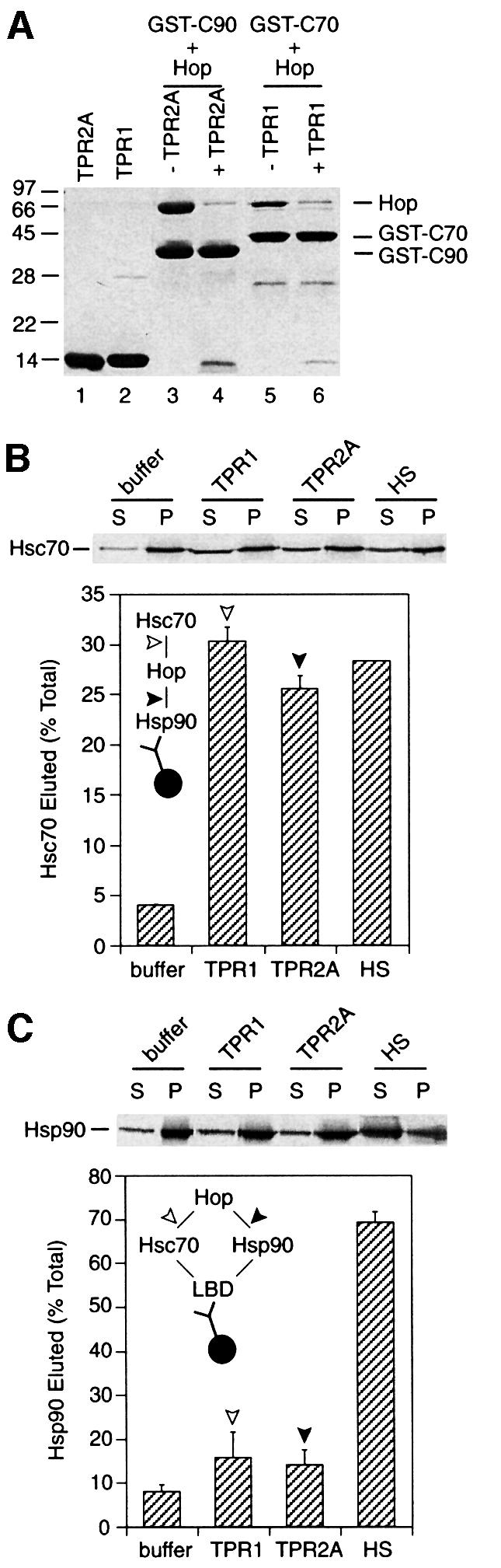
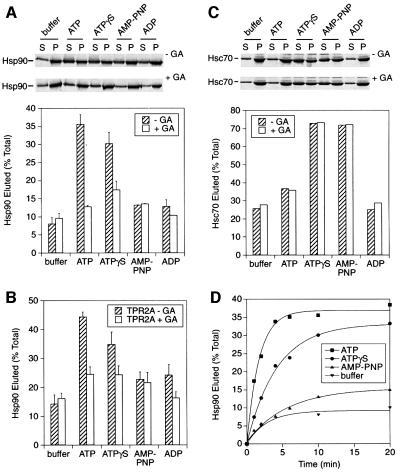
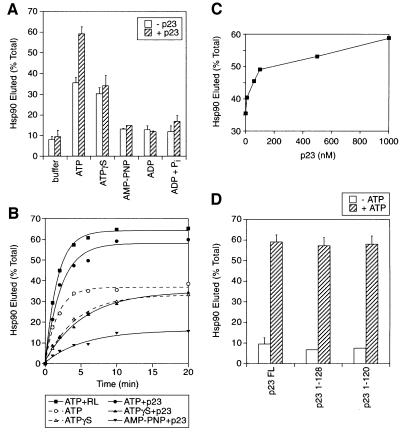
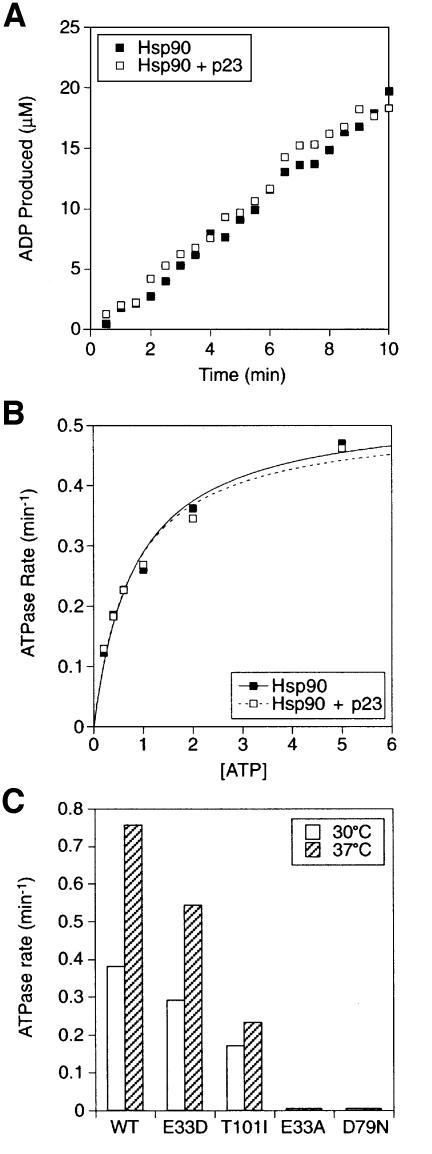
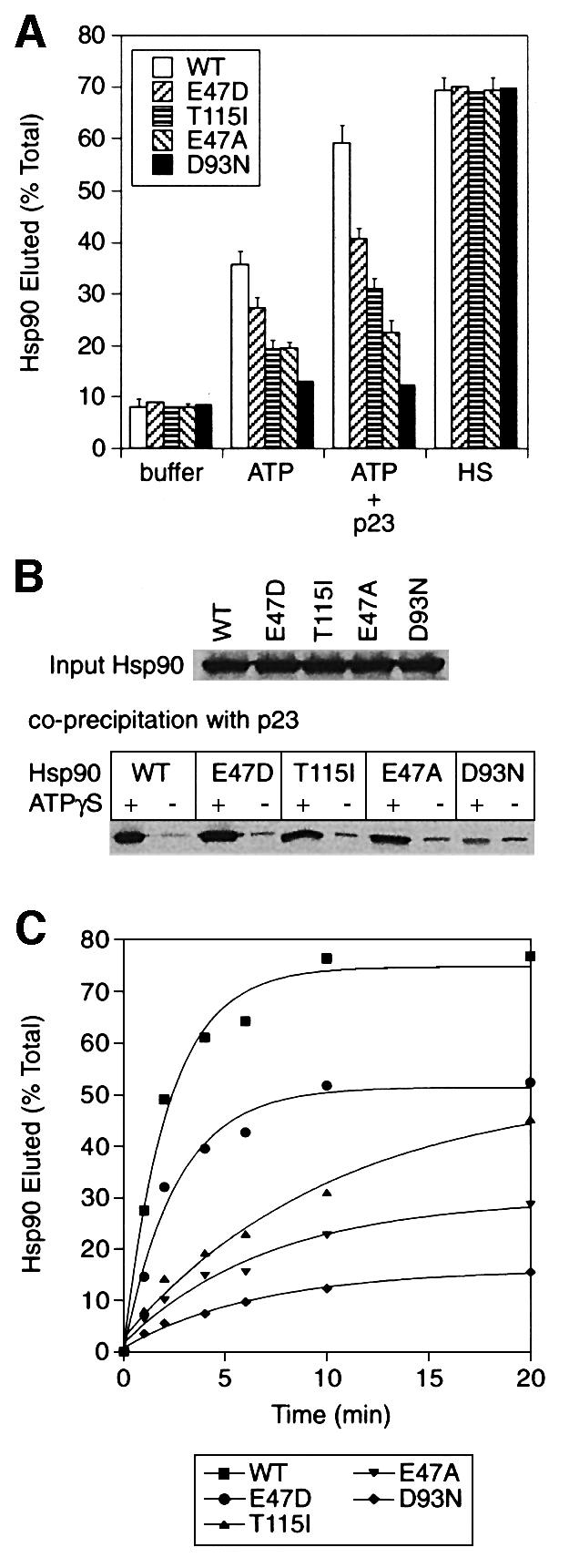
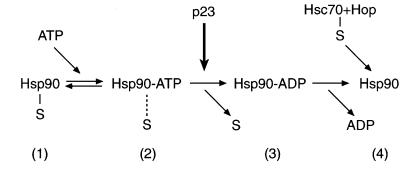
The molecular chaperone Hsp90 binds and hydrolyses ATP, but how this ATPase activity regulates the interaction of Hsp90 with a polypeptide substrate is not yet understood. Using the glucocorticoid receptor ligand binding domain as a substrate, we show that dissociation of Hsp90 from bound polypeptide depends on the Hsp90 ATPase and is blocked by geldanamycin, a specific ATPase inhibitor. The co-chaperone p23 greatly stimulates Hsp90 substrate release with ATP, but not with the non-hydrolysable nucleotides ATPgammaS or AMP-PNP. Point mutants of Hsp90 with progressively lower ATPase rates are progressively slower in ATP-dependent substrate release but are still regulated by p23. In contrast, ATPase-inactive Hsp90 mutants release substrate poorly and show no p23 effect. These results outline an ATP-driven cycle of substrate binding and release for Hsp90 which differs from that of other ATP-driven chaperones. Conversion of the ATP state of Hsp90 to the ADP state through hydrolysis is required for efficient release of substrate polypeptide. p23 couples the ATPase activity to polypeptide dissociation and thus can function as a substrate release factor for Hsp90.
Figures
Similar articles
-
Stimulation of the weak ATPase activity of human hsp90 by a client protein.J Mol Biol. 2002 Jan 25;315(4):787-98. doi: 10.1006/jmbi.2001.5245. J Mol Biol. 2002. PMID: 11812147
-
In vivo function of Hsp90 is dependent on ATP binding and ATP hydrolysis.J Cell Biol. 1998 Nov 16;143(4):901-10. doi: 10.1083/jcb.143.4.901. J Cell Biol. 1998. PMID: 9817749 Free PMC article.
-
Cofactor Tpr2 combines two TPR domains and a J domain to regulate the Hsp70/Hsp90 chaperone system.EMBO J. 2003 Jul 15;22(14):3613-23. doi: 10.1093/emboj/cdg362. EMBO J. 2003. PMID: 12853476 Free PMC article.
-
p23 and Aha1: Distinct Functions Promote Client Maturation.Subcell Biochem. 2023;101:159-187. doi: 10.1007/978-3-031-14740-1_6. Subcell Biochem. 2023. PMID: 36520307 Review.
-
Evolutionary epitopes of Hsp90 and p23: implications for their interaction.FASEB J. 2004 Jun;18(9):940-7. doi: 10.1096/fj.04-1570hyp. FASEB J. 2004. PMID: 15173105 Review.
Cited by
-
Heat shock protein 90β stabilizes focal adhesion kinase and enhances cell migration and invasion in breast cancer cells.Exp Cell Res. 2014 Aug 1;326(1):78-89. doi: 10.1016/j.yexcr.2014.05.018. Epub 2014 May 28. Exp Cell Res. 2014. PMID: 24880126 Free PMC article.
-
Novel interaction of the Hsp90 chaperone machine with Ssl2, an essential DNA helicase in Saccharomyces cerevisiae.Curr Genet. 2005 Jun;47(6):368-80. doi: 10.1007/s00294-005-0580-6. Epub 2005 May 4. Curr Genet. 2005. PMID: 15871019 Free PMC article.
-
An essential role for ATP binding and hydrolysis in the chaperone activity of GRP94 in cells.Proc Natl Acad Sci U S A. 2009 Jul 14;106(28):11600-5. doi: 10.1073/pnas.0902626106. Epub 2009 Jun 24. Proc Natl Acad Sci U S A. 2009. PMID: 19553200 Free PMC article.
-
The Role of Hsp70s in the Development and Pathogenicity of Plasmodium falciparum.Adv Exp Med Biol. 2021;1340:75-95. doi: 10.1007/978-3-030-78397-6_3. Adv Exp Med Biol. 2021. PMID: 34569021
-
Human mitochondrial import receptor Tom70 functions as a monomer.Biochem J. 2010 Aug 1;429(3):553-63. doi: 10.1042/BJ20091855. Biochem J. 2010. PMID: 20504278 Free PMC article.
References
-
- Ban C., Junop,M. and Yang,W. (1999) Transformation of MutL by ATP binding and hydrolysis: a switch in DNA mismatch repair. Cell, 97, 85–97. - PubMed
-
- Berger J.M., Gamblin,S.J., Harrison,S.C. and Wang,J.C. (1996) Structure and mechanism of DNA topoisomerase II. Nature, 379, 225–232. - PubMed
-
- Bilwes A.M., Alex,L.A., Crane,B.R. and Simon,M.I. (1999) Structure of CheA, a signal-transducing histidine kinase. Cell, 96, 131–141. - PubMed
-
- Bose S., Weikl,T., Bügl,H. and Buchner,J. (1996) Chaperone functions of hsp90-associated proteins. Science, 274, 1715–1717. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
