

DRAL is a p53-responsive gene whose four and a half LIM domain protein product induces apoptosis
- PMID: 11062252
- PMCID: PMC2185594
- DOI: 10.1083/jcb.151.3.495
DRAL is a p53-responsive gene whose four and a half LIM domain protein product induces apoptosis
Abstract
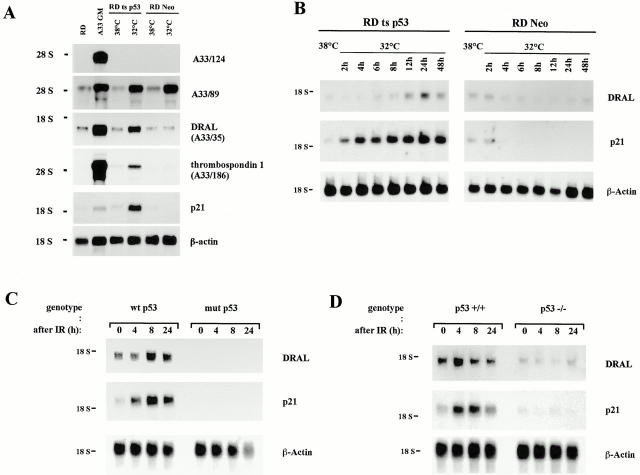
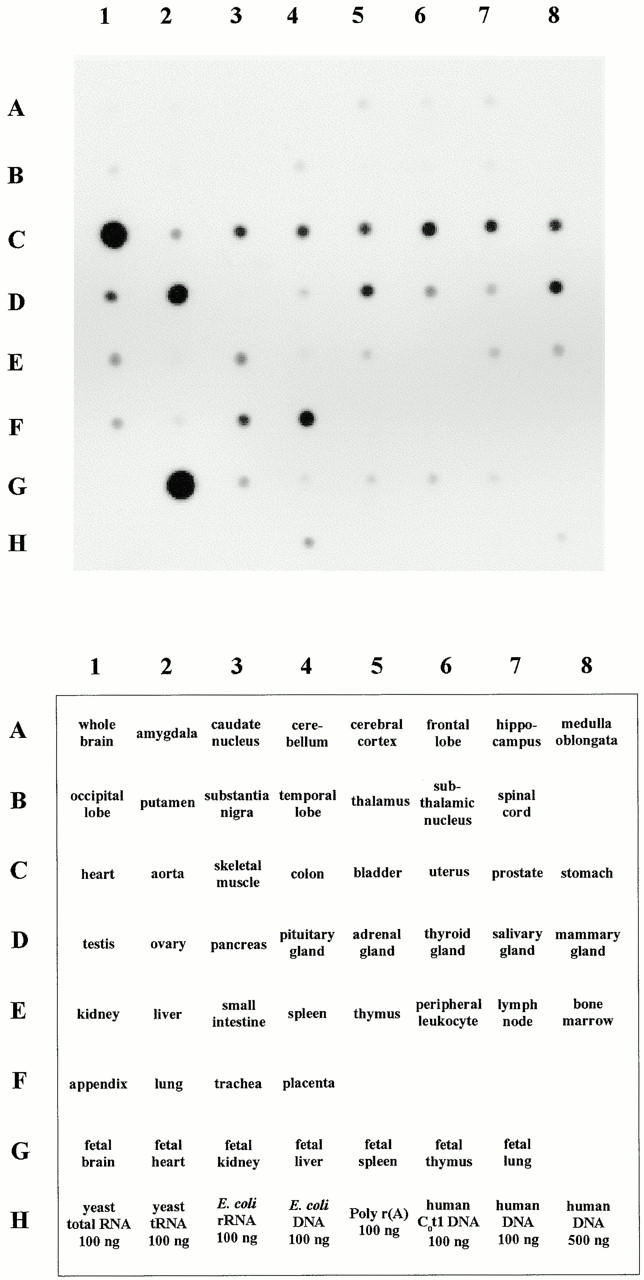
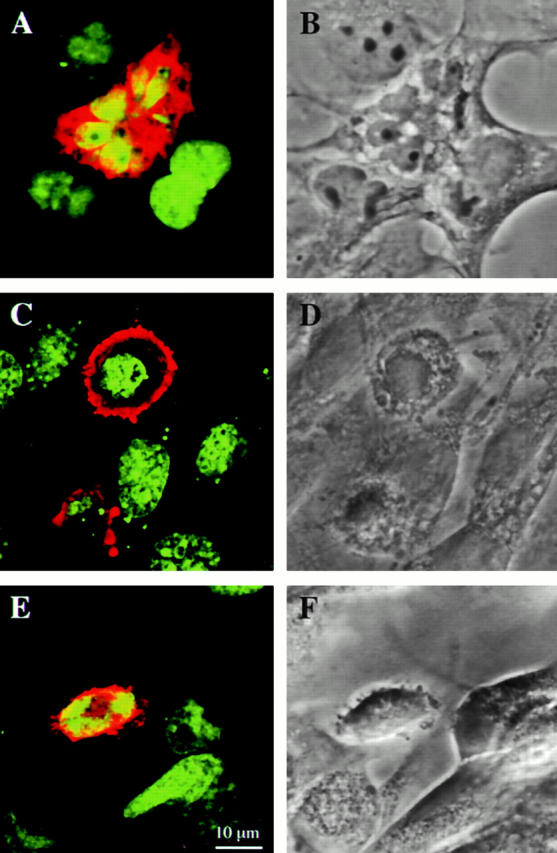
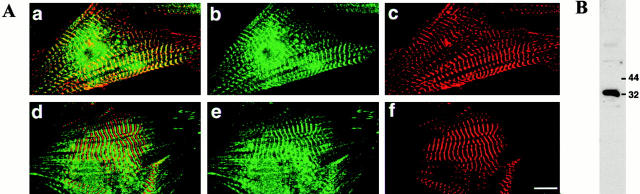
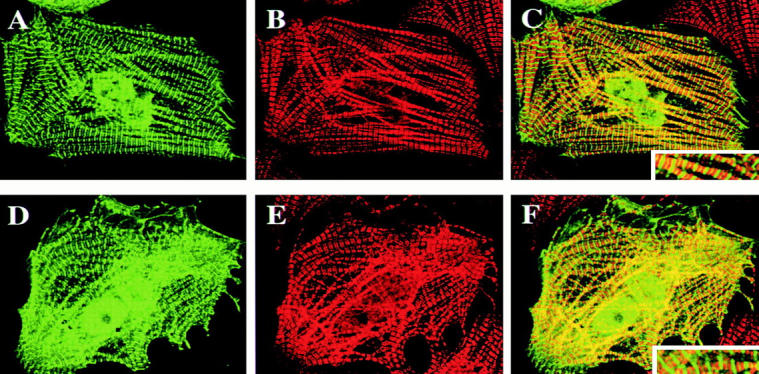
DRAL is a four and a half LIM domain protein identified because of its differential expression between normal human myoblasts and the malignant counterparts, rhabdomyosarcoma cells. In the current study, we demonstrate that transcription of the DRAL gene can be stimulated by p53, since transient expression of functional p53 in rhabdomyosarcoma cells as well as stimulation of endogenous p53 by ionizing radiation in wild-type cells enhances DRAL mRNA levels. In support of these observations, five potential p53 target sites could be identified in the promoter region of the human DRAL gene. To obtain insight into the possible functions of DRAL, ectopic expression experiments were performed. Interestingly, DRAL expression efficiently triggered apoptosis in three cell lines of different origin to the extent that no cells could be generated that stably overexpressed this protein. However, transient transfection experiments as well as immunofluorescence staining of the endogenous protein allowed for the localization of DRAL in different cellular compartments, namely cytoplasm, nucleus, focal contacts, as well as Z-discs and to a lesser extent the M-bands in cardiac myofibrils. These data suggest that downregulation of DRAL might be involved in tumor development. Furthermore, DRAL expression might be important for heart function.
Figures




References
-
- Amundson S.A., Myers T.G., Fornace A.J. Roles for p53 in growth arrest and apoptosisputting on the brakes after genotoxic stress. Oncogene. 1998;17:3287–3299. - PubMed
-
- Arber S., Hunter J.J., Ross J., Jr., Hongo M., Sansig G., Borg J., Perriard J.C., Chien K.R., Caroni P. MLP-deficient mice exhibit a disruption of cardiac cytoarchitectural organization, dilated cardiomyopathy, and heart failure. Cell. 1997;88:393–403. - PubMed
-
- Bähler M., Moser H., Eppenberger H.M., Wallimann T. Heart C-protein is transiently expressed during skeletal muscle development in the embryo, but persists in cultured myogenic cells. Dev. Biol. 1985;112:345–352. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
