

Recycling of the yeast a-factor receptor
- PMID: 11062272
- PMCID: PMC2185590
- DOI: 10.1083/jcb.151.3.731
Recycling of the yeast a-factor receptor
Abstract
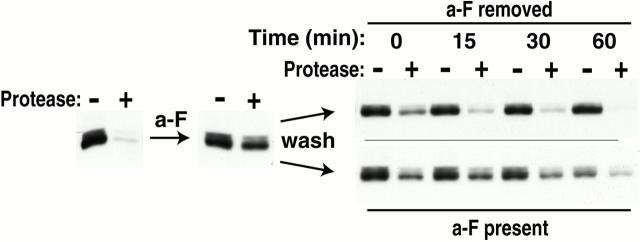
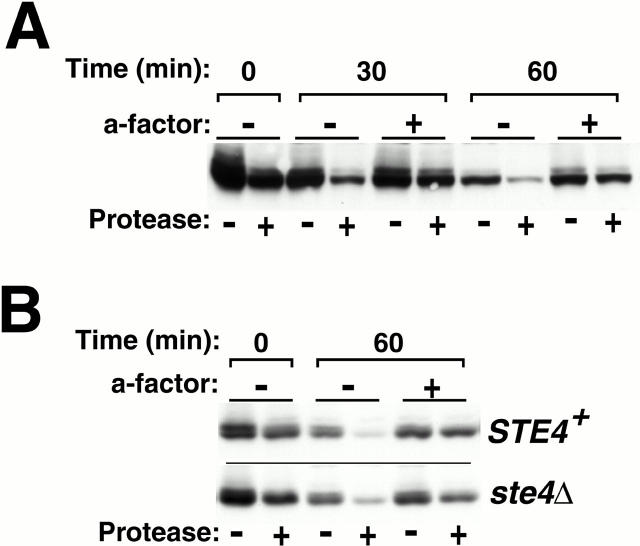
The yeast a-factor receptor (Ste3p) is subject to two mechanistically distinct modes of endocytosis: a constitutive, ligand-independent pathway and a ligand-dependent uptake pathway. Whereas the constitutive pathway leads to degradation of the receptor in the vacuole, the present work finds that receptor internalized via the ligand-dependent pathway recycles. With the a-factor ligand continuously present in the culture medium, trafficking of the receptor achieves an equilibrium in which continuing uptake to endosomal compartments is balanced by its recycling return to the plasma membrane. Withdrawal of ligand from the medium leads to a net return of the internalized receptor back to the plasma membrane. Although recycling is demonstrated for receptors that lack the signal for constitutive endocytosis, evidence is provided indicating a participation of recycling in wild-type Ste3p trafficking as well: a-factor treatment both slows wild-type receptor turnover and results in receptor redistribution to intracellular endosomal compartments. Apparently, a-factor acts as a switch, diverting receptor from vacuole-directed endocytosis and degradation, to recycling. A model is presented for how the two Ste3p endocytic modes may collaborate to generate the polarized receptor distribution characteristic of mating cells.
Figures



References
-
- Ciechanover A., Schwartz A.L., Lodish H.F. Sorting and recycling of cell surface receptors and endocytosed ligandsthe asialoglycoprotein and transferrin receptors. J. Cell Biochem. 1983;23:107–130. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
