

Nitric oxide and atrial natriuretic factor stimulate cGMP-dependent membrane insertion of aquaporin 2 in renal epithelial cells
- PMID: 11067864
- PMCID: PMC301414
- DOI: 10.1172/JCI9594
Nitric oxide and atrial natriuretic factor stimulate cGMP-dependent membrane insertion of aquaporin 2 in renal epithelial cells
Abstract
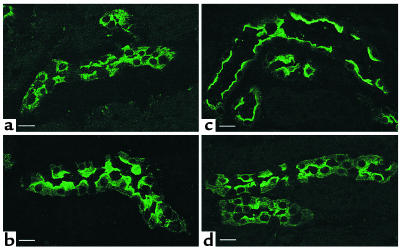
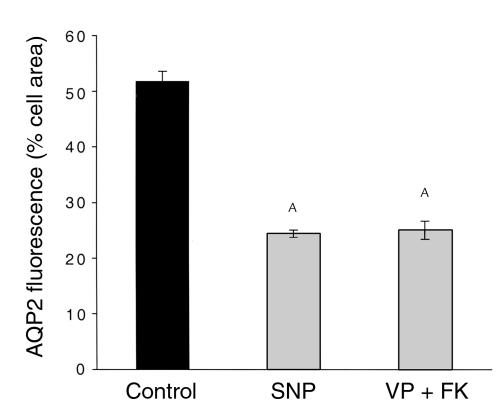
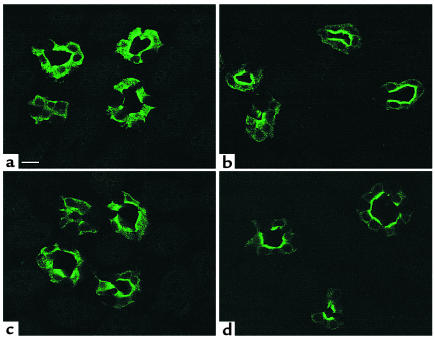
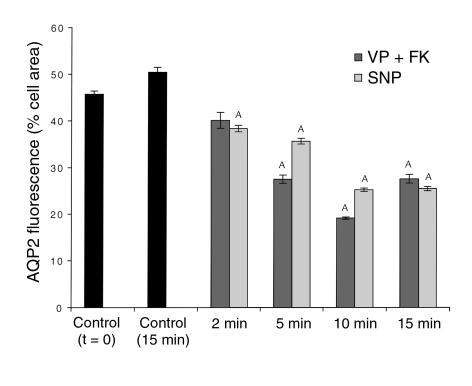
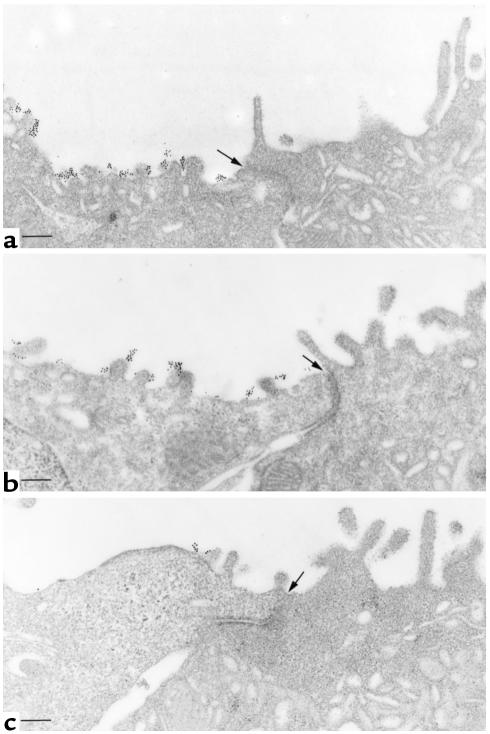
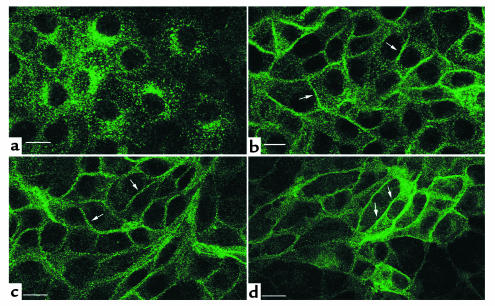
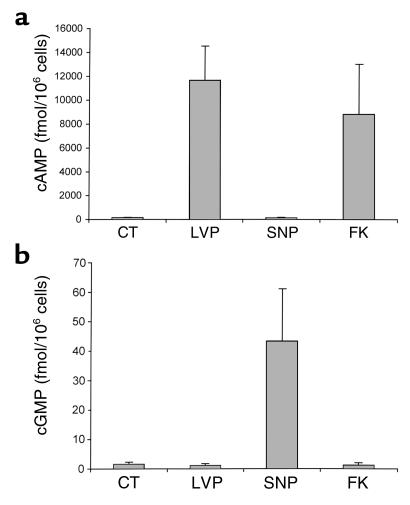
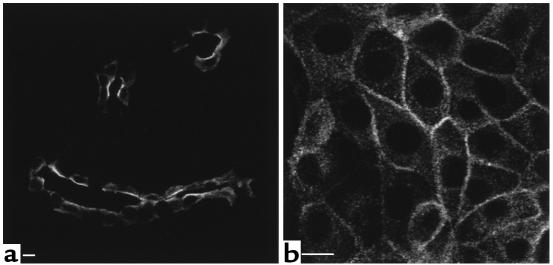
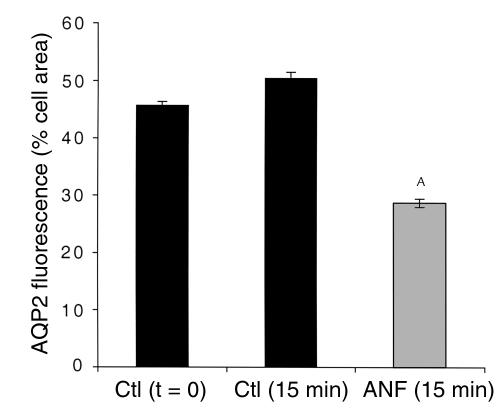
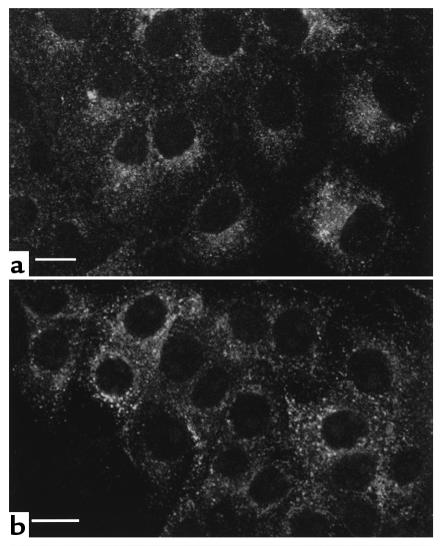
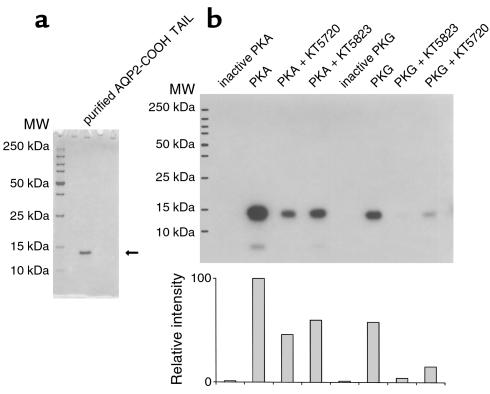
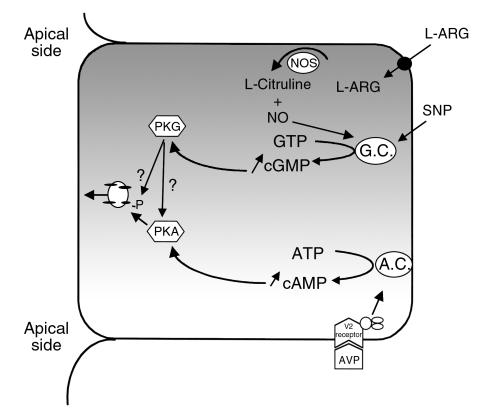
In collecting duct principal cells, aquaporin 2 (AQP2) is shuttled from intracellular vesicles to the plasma membrane upon vasopressin (VP) stimulation. VP activates adenylyl cyclase, increases intracellular cAMP, activating protein kinase A (PKA) to phosphorylate AQP2 on the COOH-terminal residue, serine 256. Using rat kidney slices and LLC-PK1 cells stably expressing AQP2 (LLC-AQP2 cells), we now show that AQP2 trafficking can be stimulated by cAMP-independent pathways. In these systems, the nitric oxide (NO) donors sodium nitroprusside (SNP) and NONOate and the NO synthase substrate L-arginine mimicked the effect of VP, stimulating relocation of AQP2 from cytoplasmic vesicles to the plasma membrane. Unlike VP, these other agents did not increase intracellular cAMP. However, SNP increased intracellular cGMP, and exogenous cGMP stimulated AQP2-membrane insertion. Atrial natriuretic factor, which signals via cGMP, also stimulated AQP2 translocation. The VP and SNP effects were blocked by the kinase inhibitor H89. SNP did not stimulate membrane insertion of AQP2 in LLC-PK1 cells expressing the phosphorylation-deficient mutant 256SerAla-AQP2, indicating that phosphorylation of Ser256 is required for signaling. Both PKA and cGMP-dependent protein kinase G phosphorylated AQP2 on this COOH-terminal residue in vitro. These results demonstrate a novel, cAMP-independent and cGMP-dependent pathway for AQP2 membrane insertion in renal epithelial cells.
Figures
References
-
- Gross SS, Wolin MS. Nitric oxide: pathophysiological mechanisms. Annu Rev Physiol. 1995;57:737–769. - PubMed
-
- Bredt DS, Snyder SH. Nitric oxide: a physiologic messenger molecule. Annu Rev Biochem. 1994;63:175–195. - PubMed
-
- Ahn KY, Mohaupt MG, Madsen KM, Kone BC. In situ hybridization localization of mRNA encoding inducible nitric oxide synthase in rat kidney. Am J Physiol. 1994;267:F748–F757. - PubMed
-
- Mohaupt MG, et al. Differential expression and induction of mRNAs encoding two inducible nitric oxide synthases in rat kidney. Kidney Int. 1994;46:653–665. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
