

Host cell invasion by Trypanosoma cruzi is potentiated by activation of bradykinin B(2) receptors
- PMID: 11067878
- PMCID: PMC2193362
- DOI: 10.1084/jem.192.9.1289
Host cell invasion by Trypanosoma cruzi is potentiated by activation of bradykinin B(2) receptors
Abstract
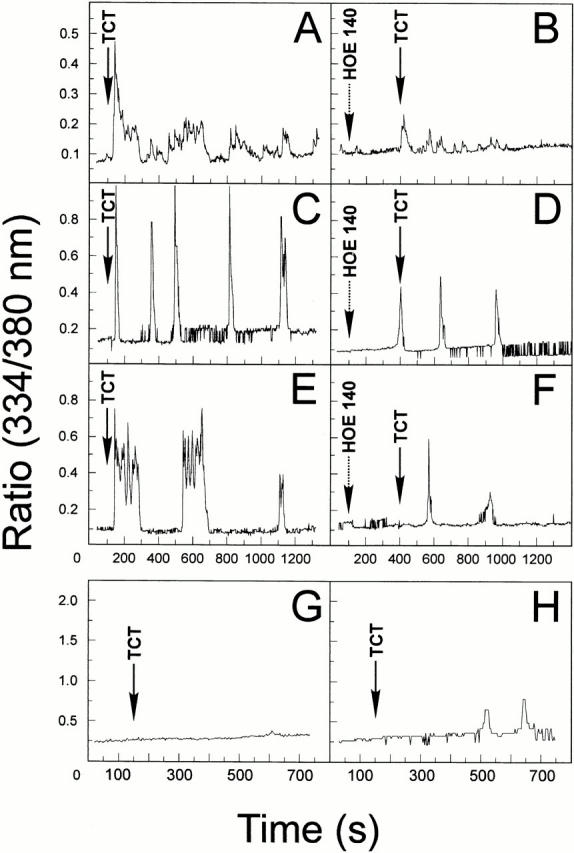
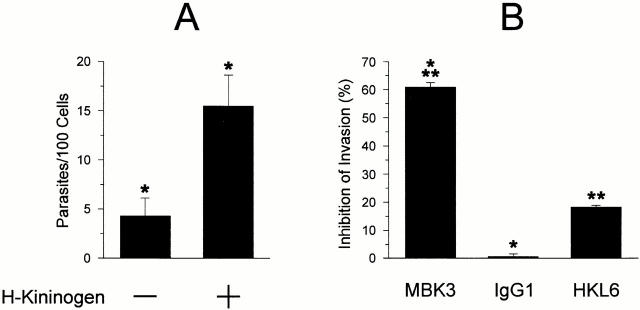
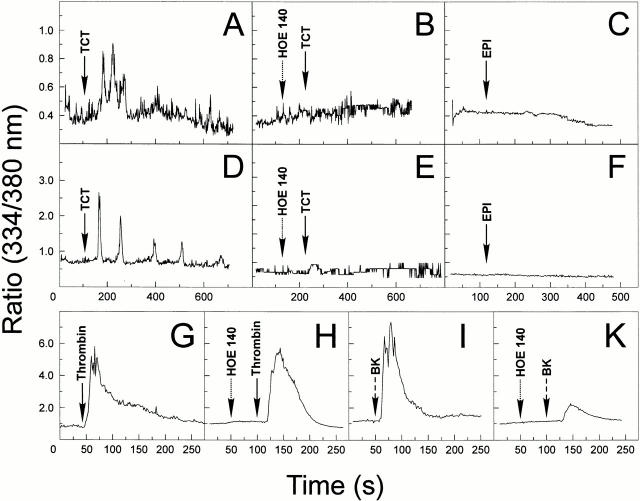
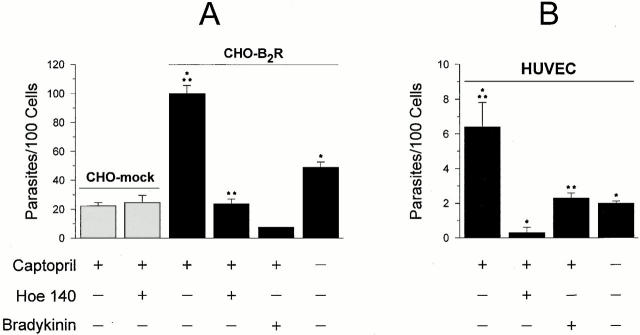
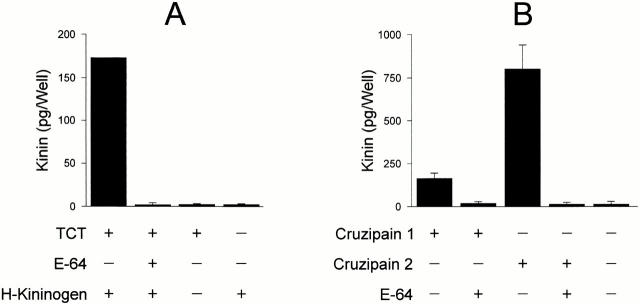
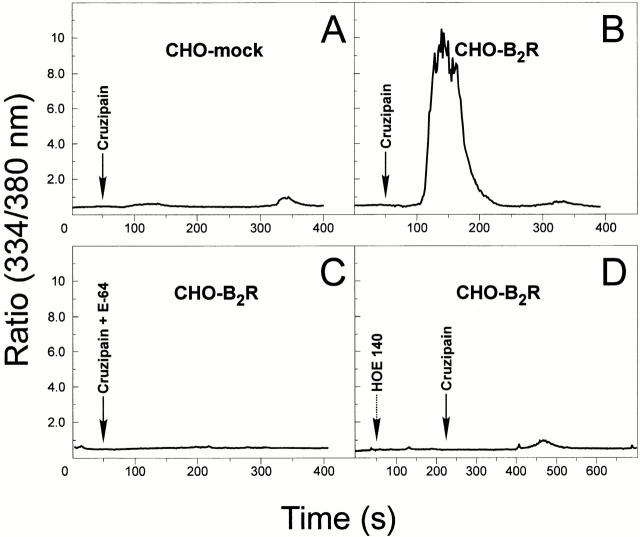
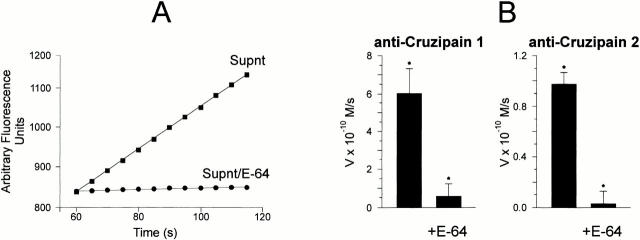
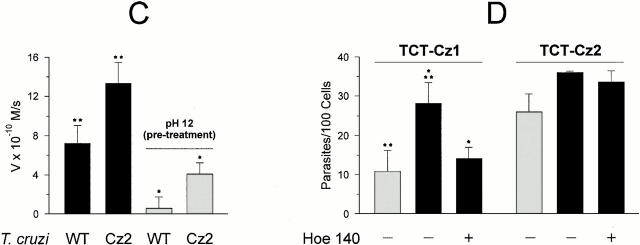
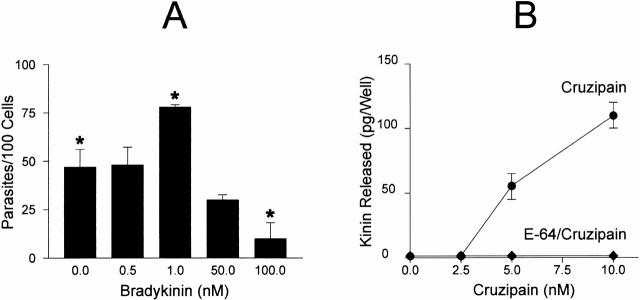
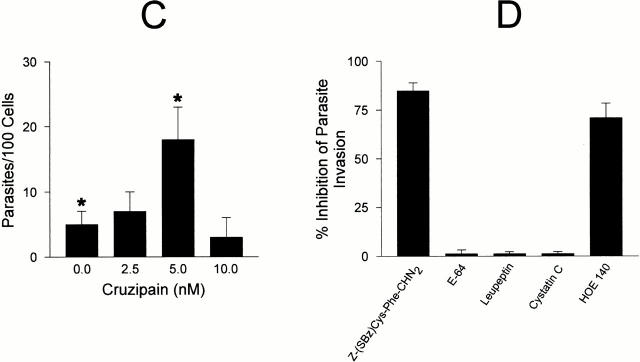
The parasitic protozoan Trypanosoma cruzi employs multiple molecular strategies to invade a broad range of nonphagocytic cells. Here we demonstrate that the invasion of human primary umbilical vein endothelial cells (HUVECs) or Chinese hamster ovary (CHO) cells overexpressing the B(2) type of bradykinin receptor (CHO-B(2)R) by tissue culture trypomastigotes is subtly modulated by the combined activities of kininogens, kininogenases, and kinin-degrading peptidases. The presence of captopril, an inhibitor of bradykinin degradation by kininase II, drastically potentiated parasitic invasion of HUVECs and CHO-B(2)R, but not of mock-transfected CHO cells, whereas the B(2)R antagonist HOE 140 or monoclonal antibody MBK3 to bradykinin blocked these effects. Invasion competence correlated with the parasites' ability to liberate the short-lived kinins from cell-bound kininogen and to elicit vigorous intracellular free calcium ([Ca(2+)](i)) transients through B(2)R. Invasion was impaired by membrane-permeable cysteine proteinase inhibitors such as Z-(SBz)Cys-Phe-CHN(2) but not by the hydrophilic inhibitor 1-trans-epoxysuccinyl-l-leucyl-amido-(4-guanidino) butane or cystatin C, suggesting that kinin release is confined to secluded spaces formed by juxtaposition of host cell and parasite plasma membranes. Analysis of trypomastigote transfectants expressing various cysteine proteinase isoforms showed that invasion competence is linked to the kinin releasing activity of cruzipain, herein proposed as a factor of virulence in Chagas' disease.
Figures
References
-
- Andrade Z.A. Immunopathology of Chagas' disease Mem. Inst. Oswaldo Cruz. 94Suppl. I1999. 71 80 - PubMed
-
- Rossi M. Microvascular changes as a cause of chronic cardiomyopathy in Chagas' disease. Am. Heart J. 1990;120:233–236. - PubMed
-
- Morris S.A., Tanowitz H., Wittner M., Bilezikjan J.P. Pathophysiological insights into the cardiomyopathy of Chagas' disease. Circulation. 1990;82:1900–1909. - PubMed
-
- Tardieux I., Webster P., Ravesloot J., Boron W., Lunn J.A., Andrews N.W. Lysosome recruitment and fusion are early events required for Trypanosoma invasion of mammalian cells. Cell. 1992;71:1117–1130. - PubMed
-
- Burleigh B.A, Andrews N.W. The mechanisms of Trypanosoma cruzi invasion of mammalian cells. Annu. Rev. Microbiol. 1995;49:175–200. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
