

Pax6 activity in the lens primordium is required for lens formation and for correct placement of a single retina in the eye
- PMID: 11069887
- PMCID: PMC317031
- DOI: 10.1101/gad.184000
Pax6 activity in the lens primordium is required for lens formation and for correct placement of a single retina in the eye
Abstract
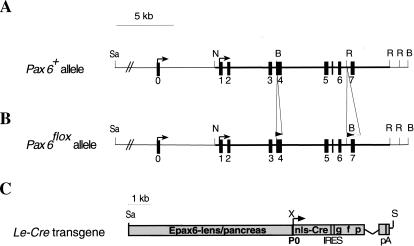
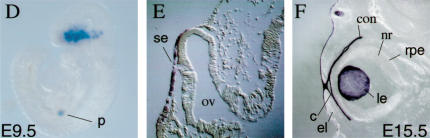
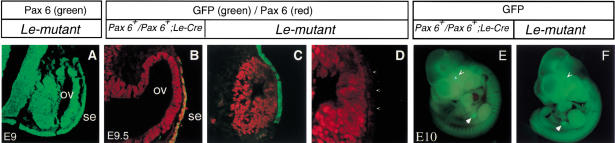
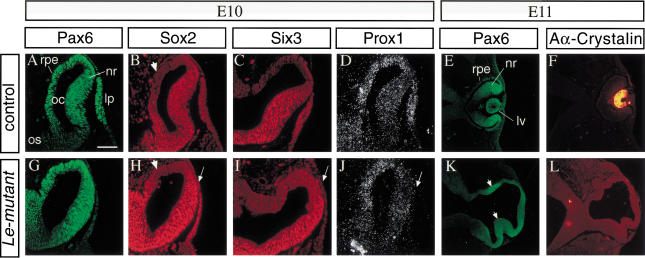
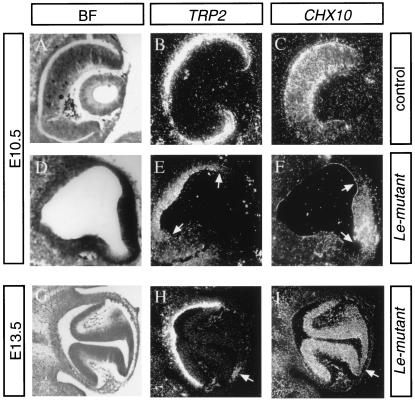
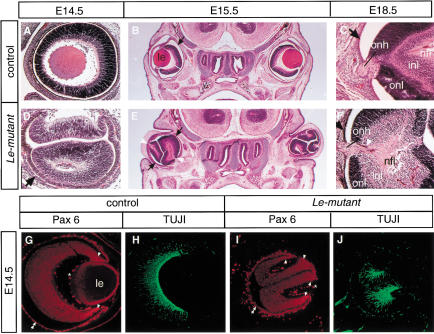
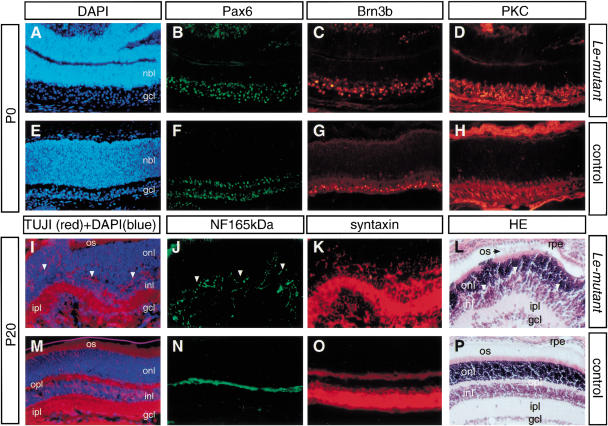
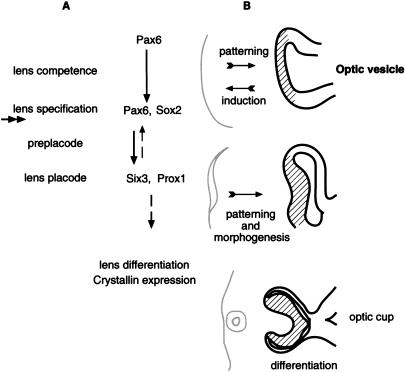
The Pax6 transcription factor plays a key role in ocular development of vertebrates and invertebrates. Homozygosity of the Pax6 null mutation in human and mice results in arrest of optic vesicle development and failure to initiate lens formation. This phenotype obscures the understanding of autonomous function of Pax6 in these tissue components and during later developmental stages. We employed the Cre/loxP approach to inactivate Pax6 specifically in the eye surface ectoderm concomitantly with lens induction. Although lens induction occurred in the mutant, as indicated by Sox2 up-regulation in the surface ectoderm, further development of the lens was arrested. Hence, Pax6 activity was found to be essential in the specified ectoderm for lens placode formation. Furthermore, this mutant model allowed us for the first time to address in vivo the development of a completely normal retina in the absence of early lens structures. Remarkably, several independent, fully differentiated neuroretinas developed in a single optic vesicle in the absence of a lens, demonstrating that the developing lens is not necessary to instruct the differentiation of the neuroretina but is, rather, required for the correct placement of a single retina in the eye.
Figures
References
-
- Barnstable CJ. A molecular view of vertebrate retinal development. Mol Neurobiol. 1987;1:9–46. - PubMed
-
- Collinson JM, Hill RE, West JD. Different roles for pax6 in the optic vesicle and facial epithelium mediate early morphogenesis of the murine eye. Development. 2000;127:945–956. - PubMed
-
- Coulombre AJ, Coulombre JL. Lens development. I. Role of the lens in eye growth. J Exp Zool. 1964;156:39–47. - PubMed
-
- Cvekl A, Piatigorsky J. Lens development and crystallin gene expression: Many roles for Pax-6. Bioessays. 1996;18:621–630. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials