

Phosphorylation status of the parvovirus minute virus of mice particle: mapping and biological relevance of the major phosphorylation sites
- PMID: 11069983
- PMCID: PMC113168
- DOI: 10.1128/jvi.74.23.10892-10902.2000
Phosphorylation status of the parvovirus minute virus of mice particle: mapping and biological relevance of the major phosphorylation sites
Abstract
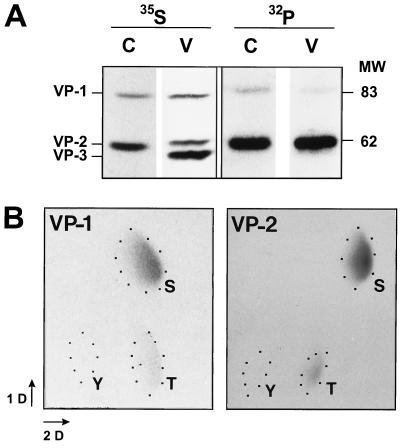
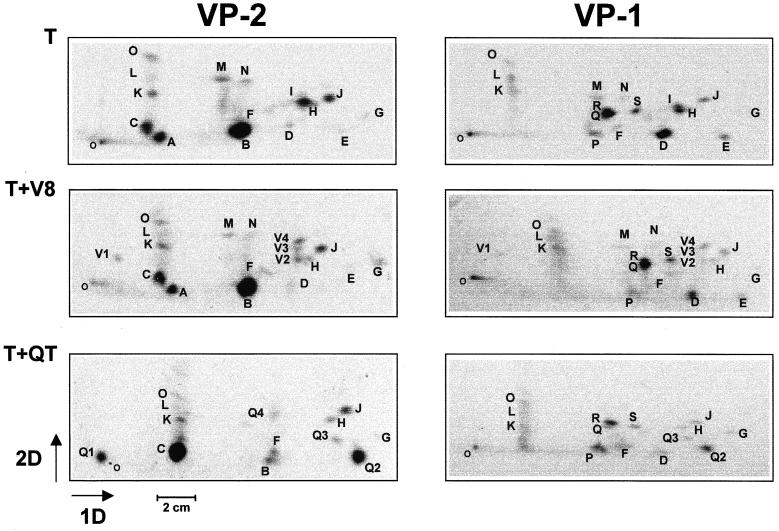
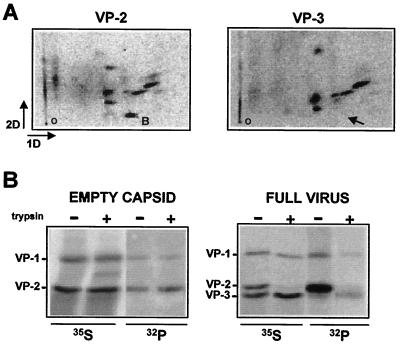
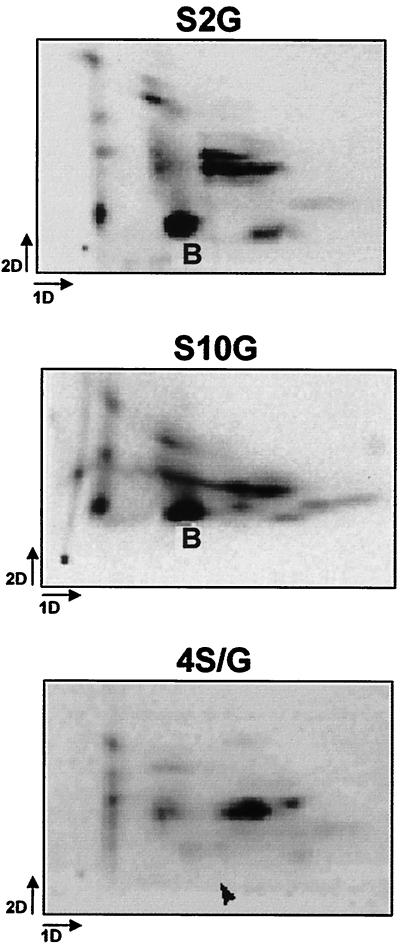
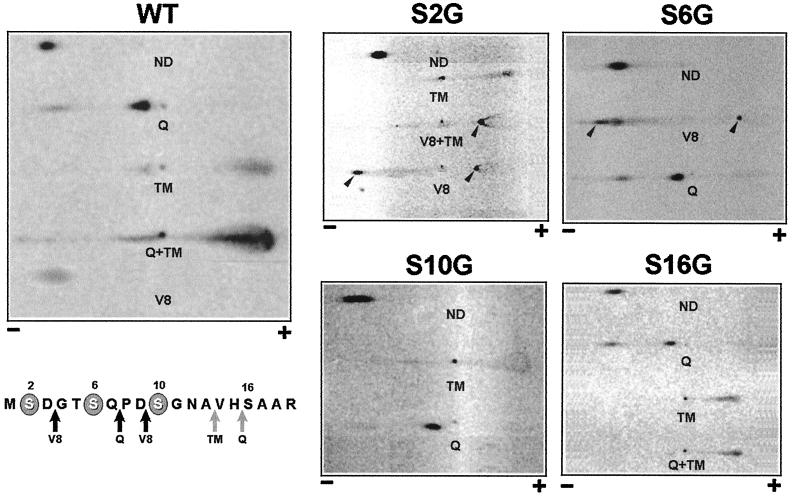
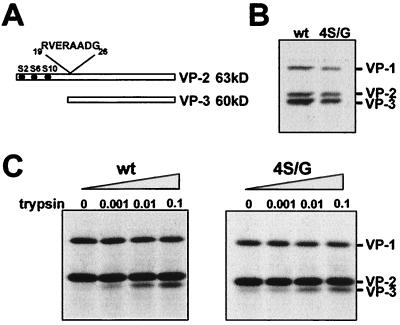
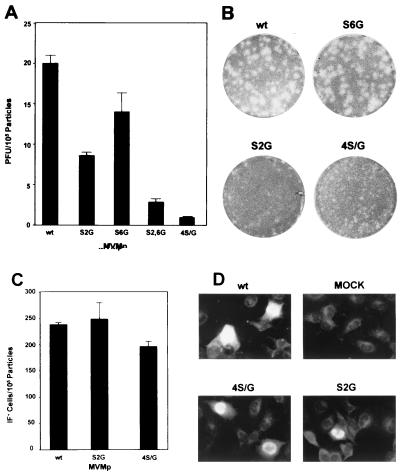
The core of the VP-1 and VP-2 proteins forming the T=1 icosahedral capsid of the prototype strain of the parvovirus minute virus of mice (MVMp) share amino acids sequence and a common three-dimensional structure; however, the roles of these polypeptides in the virus infection cycle differ. To gain insights into this paradox, the nature, distribution, and biological significance of MVMp particle phosphorylation was investigated. The VP-1 and VP-2 proteins isolated from purified empty capsids and from virions containing DNA harbored phosphoserine and phosphothreonine amino acids, which in two-dimensional tryptic analysis resulted in complex patterns reproducibly composed by more than 15 unevenly phosphorylated peptides. Whereas secondary protease digestions and comigration of most weak peptides in the fingerprints revealed common phosphorylation sites in the VP-1 and VP-2 subunits assembled in capsids, the major tryptic phosphopeptides were remarkably characteristic of either polypeptide. The VP-2-specific peptide named B, containing the bulk of the (32)P label of the MVMp particle in the form of phosphoserine, was mapped to the structurally unordered N-terminal domain of this polypeptide. Mutations in any or all four serine residues present in peptide B showed that the VP-2 N-terminal domain is phosphorylated at multiple sites, even though none of them was essential for capsid assembly or virus formation. Chromatographic analysis of purified wild-type (wt) and mutant peptide B digested with a panel of specific proteases allowed us to identify the VP-2 residues Ser-2, Ser-6, and Ser-10 as the main phosphate acceptors for MVMp capsid during the natural viral infection. Phosphorylation at VP-2 N-terminal serines was not necessary for the externalization of this domain outside of the capsid shell in particles containing DNA. However, the plaque-forming capacity and plaque size of VP-2 N-terminal phosphorylation mutants were severely reduced, with the evolutionarily conserved Ser-2 determining most of the phenotypic effect. In addition, the phosphorylated amino acids were not required for infection initiation or for nuclear translocation of the expressed structural proteins, and thus a role at a late stage of MVMp life cycle is proposed. This study illustrates the complexity of posttranslational modification of icosahedral viral capsids and underscores phosphorylation as a versatile mechanism to modulate the biological functions of their protein subunits.
Figures

Similar articles
-
Nuclear export of the nonenveloped parvovirus virion is directed by an unordered protein signal exposed on the capsid surface.J Virol. 2004 Oct;78(19):10685-94. doi: 10.1128/JVI.78.19.10685-10694.2004. J Virol. 2004. PMID: 15367635 Free PMC article.
-
Protein species of the parvovirus minute virus of mice strain MVMp: involvement of phosphorylated VP-2 subtypes in viral morphogenesis.J Virol. 1993 Sep;67(9):5126-38. doi: 10.1128/JVI.67.9.5126-5138.1993. J Virol. 1993. PMID: 8394440 Free PMC article.
-
Biochemical and physical characterization of parvovirus minute virus of mice virus-like particles.Virology. 2000 Feb 15;267(2):299-309. doi: 10.1006/viro.1999.0123. Virology. 2000. PMID: 10662625
-
A beta-stranded motif drives capsid protein oligomers of the parvovirus minute virus of mice into the nucleus for viral assembly.J Virol. 2000 Apr;74(8):3804-14. doi: 10.1128/jvi.74.8.3804-3814.2000. J Virol. 2000. PMID: 10729155 Free PMC article.
-
Complementary roles of multiple nuclear targeting signals in the capsid proteins of the parvovirus minute virus of mice during assembly and onset of infection.J Virol. 2002 Jul;76(14):7049-59. doi: 10.1128/jvi.76.14.7049-7059.2002. J Virol. 2002. PMID: 12072505 Free PMC article.
Cited by
-
Post-translational modifications in capsid proteins of recombinant adeno-associated virus (AAV) 1-rh10 serotypes.FEBS J. 2019 Dec;286(24):4964-4981. doi: 10.1111/febs.15013. Epub 2019 Aug 1. FEBS J. 2019. PMID: 31330090 Free PMC article.
-
Nuclear export of the nonenveloped parvovirus virion is directed by an unordered protein signal exposed on the capsid surface.J Virol. 2004 Oct;78(19):10685-94. doi: 10.1128/JVI.78.19.10685-10694.2004. J Virol. 2004. PMID: 15367635 Free PMC article.
-
Low pH-dependent endosomal processing of the incoming parvovirus minute virus of mice virion leads to externalization of the VP1 N-terminal sequence (N-VP1), N-VP2 cleavage, and uncoating of the full-length genome.J Virol. 2006 Jan;80(2):1015-24. doi: 10.1128/JVI.80.2.1015-1024.2006. J Virol. 2006. PMID: 16379002 Free PMC article.
-
Assembly of Structurally Simple Icosahedral Viruses.Subcell Biochem. 2024;105:403-430. doi: 10.1007/978-3-031-65187-8_11. Subcell Biochem. 2024. PMID: 39738953 Review.
-
Viral oncolysis that targets Raf-1 signaling control of nuclear transport.J Virol. 2010 Feb;84(4):2090-9. doi: 10.1128/JVI.01550-09. Epub 2009 Nov 25. J Virol. 2010. PMID: 19939915 Free PMC article.
References
-
- Agbandje M, McKenna R, Rossmann M G, Strassheim M L, Parrish C R. Structure determination of feline panleukopenia virus empty particles. Proteins. 1993;16:155–171. - PubMed
-
- Agbandje M, Kajigaya S, McKenna R, Young N S, Rossmann M G. The structure of human parvovirus B19 at 8 Å resolution. Virology. 1994;203:106–115. - PubMed
-
- Agbandje-McKenna M, Llamas-Sainz A L, Wang F, Tattersall P, Rossmann M G. Functional implications of the structure of the murine parvovirus, minute virus of mice. Structure. 1998;6:1369–1381. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
