

Respiratory syncytial virus can tolerate an intergenic sequence of at least 160 nucleotides with little effect on transcription or replication in vitro and in vivo
- PMID: 11069997
- PMCID: PMC113182
- DOI: 10.1128/jvi.74.23.11017-11026.2000
Respiratory syncytial virus can tolerate an intergenic sequence of at least 160 nucleotides with little effect on transcription or replication in vitro and in vivo
Abstract
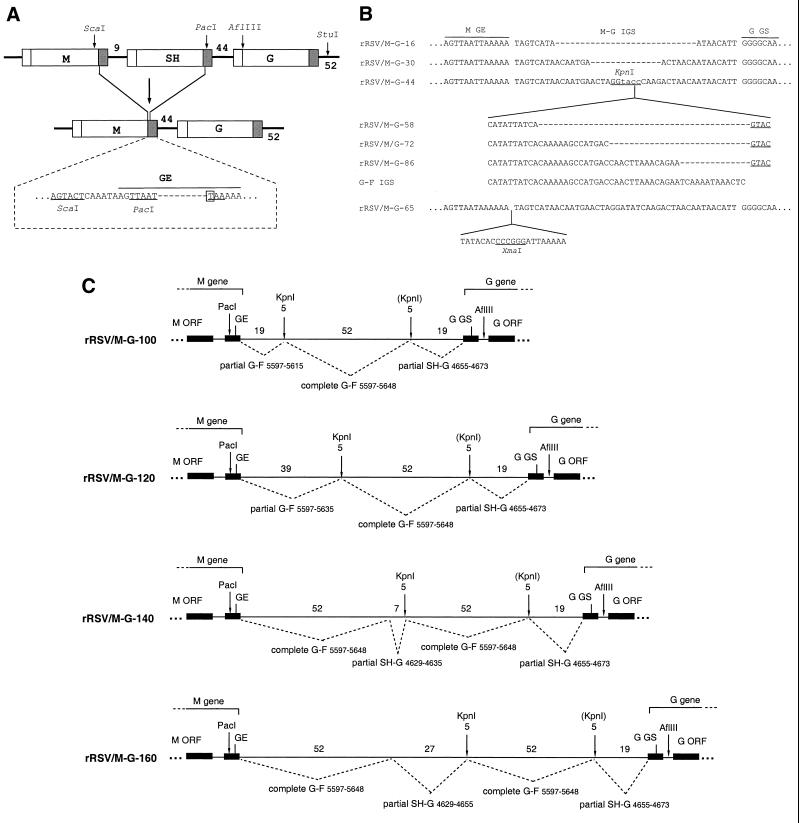
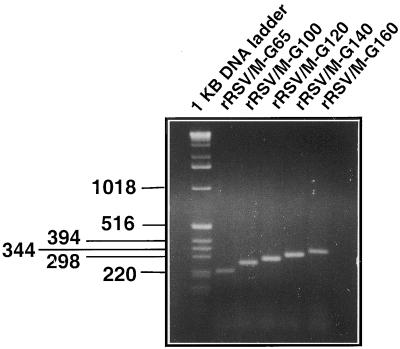
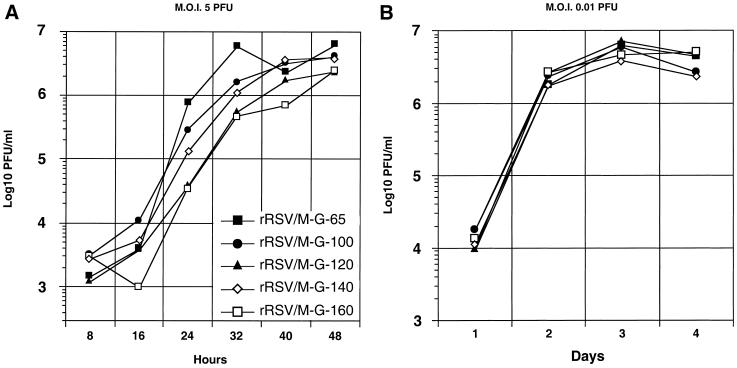
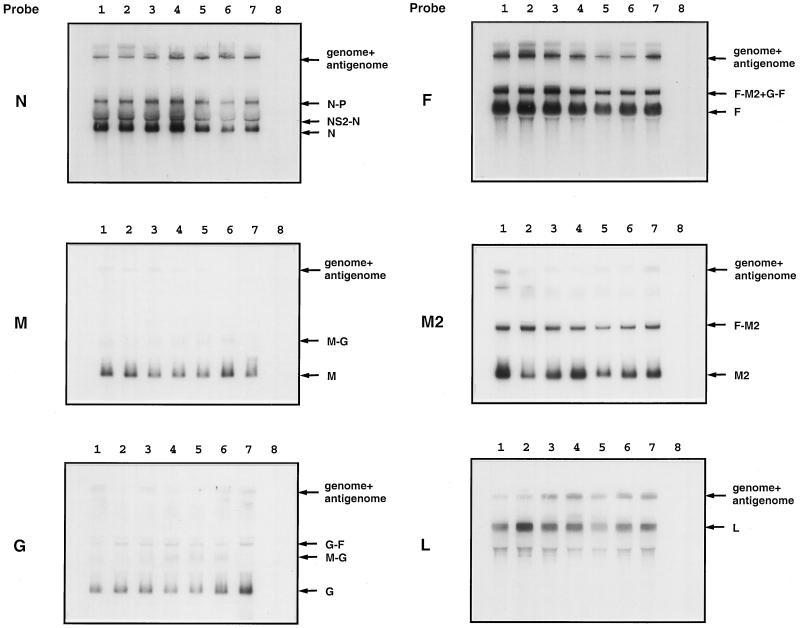
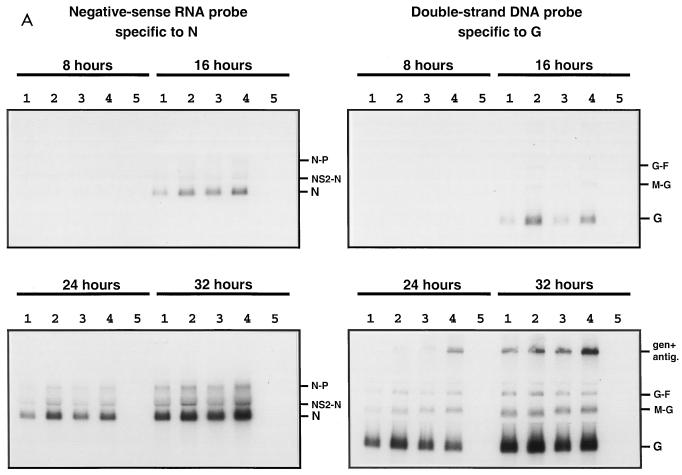
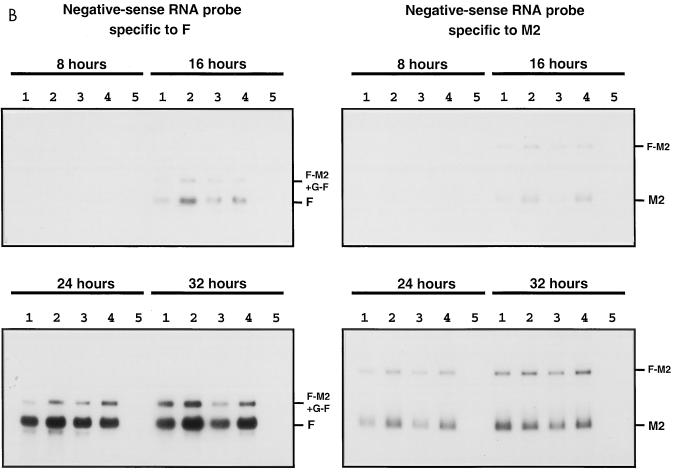
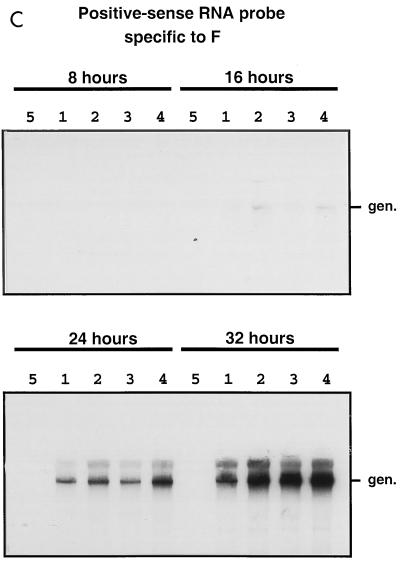
The intergenic sequences (IGS) between the first nine genes of human respiratory syncytial virus (RSV) vary in length from 1 to 56 nucleotides and lack apparent conserved sequence motifs. To investigate their influence on sequential transcription and viral growth, recombinant RSV strain A2, from which the SH gene had been deleted to facilitate manipulation, was further modified to contain an M-G IGS of 16, 30, 44, 58, 65, 72, 86, 100, 120, 140, or 160 nucleotides. All of the viruses were viable. For viruses with an M-G IGS of 100 nucleotides or more, plaque size decreased with increasing IGS length. In this same length range, increasing IGS length was associated with modest attenuation during single-step, but not multistep, growth in HEp-2 cells. Surprisingly, Northern blot analysis of the accumulation of six different mRNAs indicated that there was little or no change in transcription with increasing IGS length. Thus, the RSV polymerase apparently can readily cross IGS of various lengths, including unnaturally long ones, with little or no effect on the efficiency of termination and reinitiation. This finding supports the view that the IGS do not have much effect on sequential transcription and provides evidence from infectious virus that IGS length is not an important regulatory feature. To evaluate replication in vivo, BALB/c mice were infected intranasally with RSV containing an M-G IGS of 65, 140, or 160 nucleotides. Replication of the latter two viruses was decreased up to 5- and 25-fold in the upper and lower respiratory tracts, respectively, on day 3 following infection. However, the level of replication at both sites on days 4 and 5 was very similar to that of the virus with an IGS of 65 nucleotides. Thus, the modest attenuation in vivo associated with the longer IGS was additive to that conferred by deletion of the SH gene and might be useful to incrementally increase the level of attenuation of a live-attenuated vaccine virus.
Figures
References
-
- Collins P L, Dickens L E, Buckler-White A, Olmsted R A, Spriggs M K, Camargo E, Coelingh K V. Nucleotide sequences for the gene junctions of human respiratory syncytial virus reveal distinctive features of intergenic structure and gene order. Proc Natl Acad Sci USA. 1986;83:4594–4598. - PMC - PubMed
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
