

Efficient and specific initiation of subgenomic RNA synthesis by cucumber mosaic virus replicase in vitro requires an upstream RNA stem-loop
- PMID: 11070017
- PMCID: PMC113212
- DOI: 10.1128/jvi.74.23.11201-11209.2000
Efficient and specific initiation of subgenomic RNA synthesis by cucumber mosaic virus replicase in vitro requires an upstream RNA stem-loop
Abstract
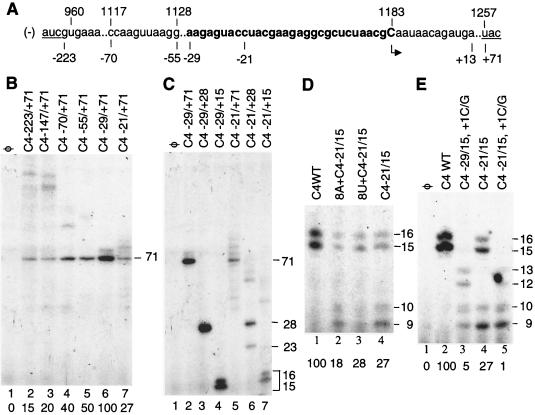
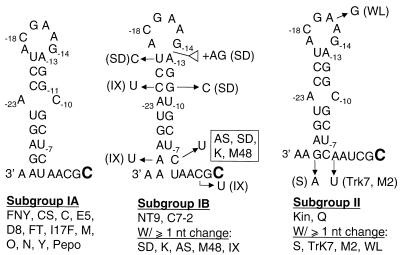
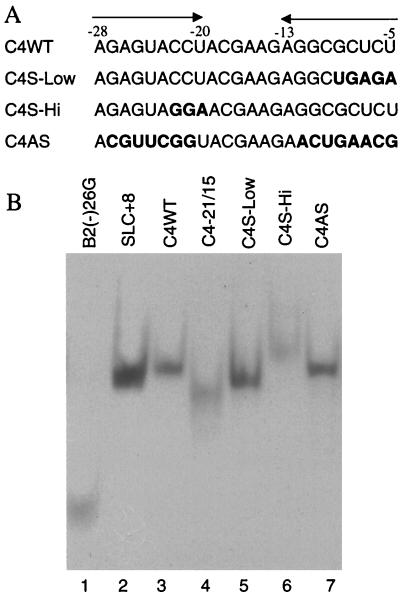
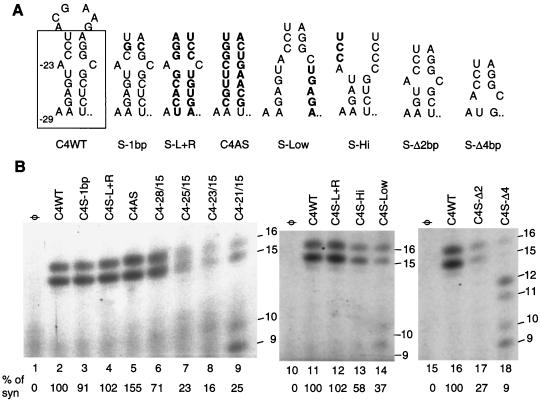
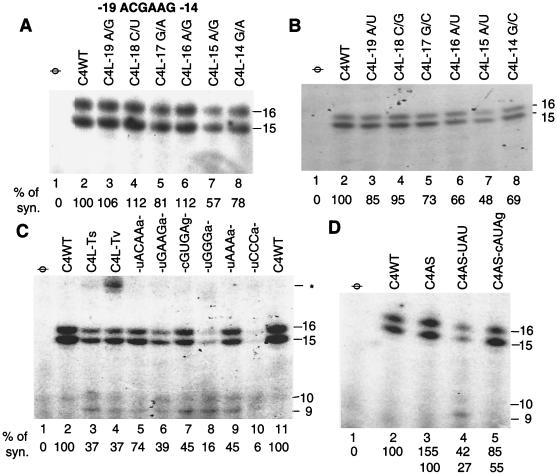
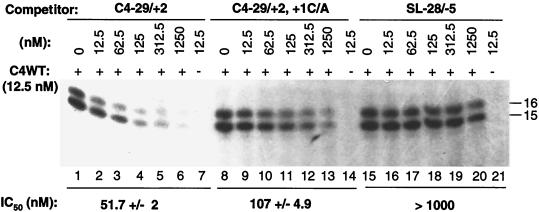
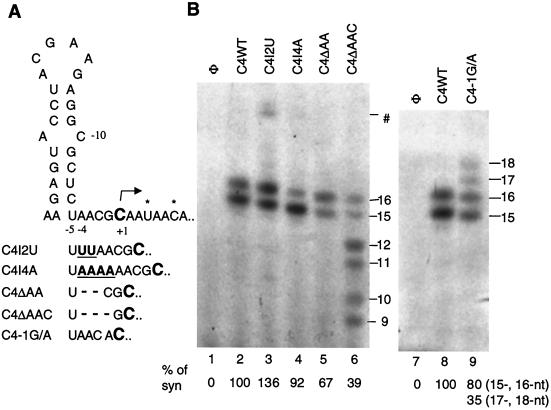
We defined the minimal core promoter sequences responsible for efficient and accurate initiation of cucumber mosaic virus (CMV) subgenomic RNA4. The necessary sequence maps to positions -28 to +15 relative to the initiation cytidylate used to initiate RNA synthesis in vivo. Positions -28 to -5 contain a 9-bp stem and a 6-nucleotide purine-rich loop. Considerable changes in the stem and the loop are tolerated for RNA synthesis, including replacement with a different stem-loop. In a template competition assay, the stem-loop and the initiation cytidylate are sufficient to interact with the CMV replicase. Thus, the mechanism of core promoter recognition by the CMV replicase appears to be less specific in comparison to the minimal subgenomic core promoter of the closely related brome mosaic virus.
Figures
References
-
- Adkins S, Kao C C. Subgenomic RNA promoters dictate the mode of recognition by bromoviral RNA-dependent RNA polymerases. Virology. 1998;252:1–8. - PubMed
-
- Boccard F, Baulcombe D. Mutational analysis of cis-acting sequences and gene function in RNA3 of cucumber mosaic virus. Virology. 1993;193:563–578. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
