

Constitutive instability of muscle regulatory factor Myf5 is distinct from its mitosis-specific disappearance, which requires a D-box-like motif overlapping the basic domain
- PMID: 11073992
- PMCID: PMC86547
- DOI: 10.1128/MCB.20.23.8923-8932.2000
Constitutive instability of muscle regulatory factor Myf5 is distinct from its mitosis-specific disappearance, which requires a D-box-like motif overlapping the basic domain
Abstract
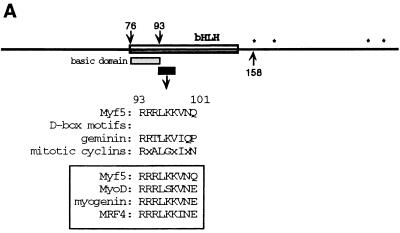
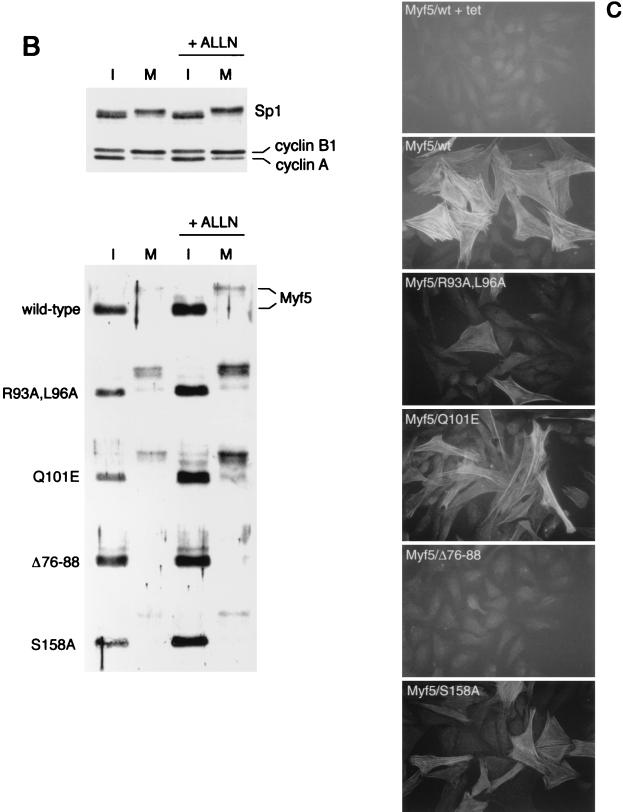
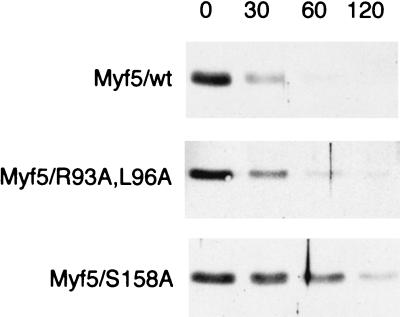
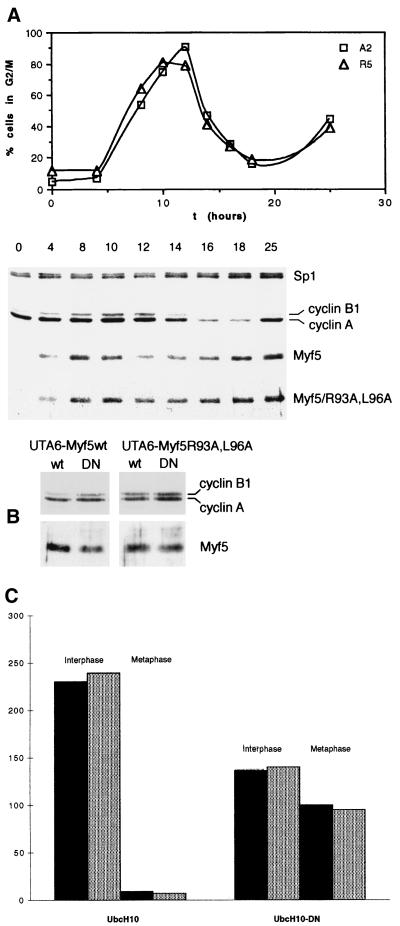
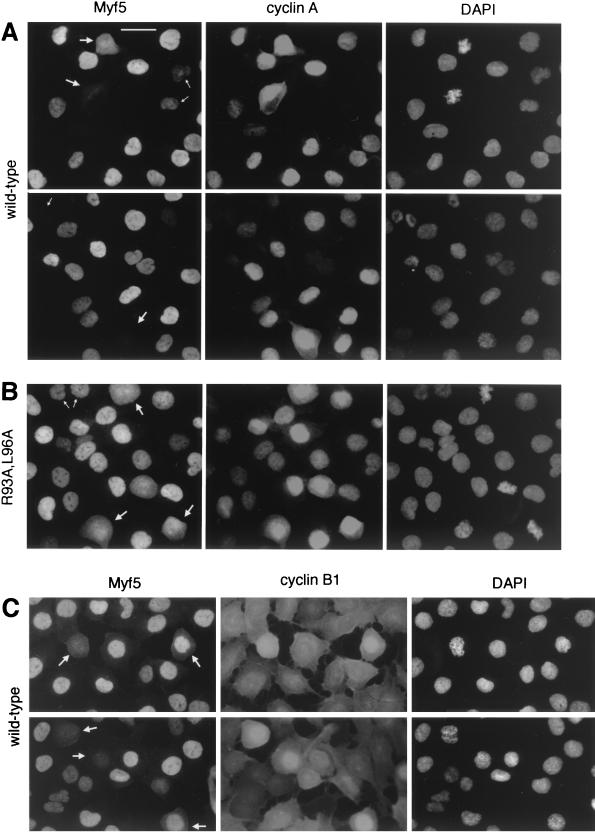
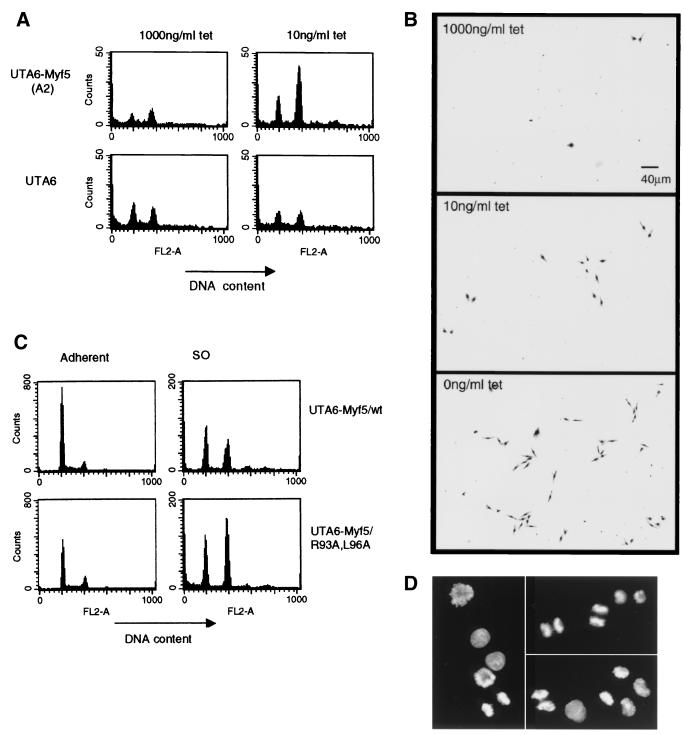
Transcription factors Myf5 and MyoD play critical roles in controlling myoblast identity and differentiation. In the myogenic cell line C2, we have found that Myf5 expression, unlike that of MyoD, is restricted to cycling cells and regulated by proteolysis at mitosis. In the present study, we have examined Myf5 proteolysis through stable transfection of myogenically convertible U20S cells with Myf5 derivatives under the control of a tetracycline-sensitive promoter. A motif within the basic helix-loop-helix domain of Myf5 (R93 to Q101) resembles the "destruction box" characteristic of substrates of mitotic proteolysis and thought to be recognized by the anaphase-promoting complex or cyclosome (APC). Mutation of this motif in Myf5 stabilizes the protein at mitosis but does not affect its constitutive turnover. Conversely, mutation of a serine residue (S158) stabilizes Myf5 in nonsynchronized cultures but not at mitosis. Thus, at least two proteolytic pathways control Myf5 levels in cycling cells. The mitotic proteolysis of Myf5 is unlike that which has been described for other destruction box-dependent substrates: down-regulation of Myf5 at mitosis appears to precede that of known targets of the APC and is not affected by a dominant-negative version of the ubiquitin carrier protein UbcH10, implicated in the APC-mediated pathway. Finally, we find that induction of Myf5 perturbs the passage of cells through mitosis, suggesting that regulation of Myf5 levels at mitosis may influence cell cycle progression of Myf5-expressing muscle precursor cells.
Figures
References
-
- Albagli O, Lantoine D, Quief S, Quignon F, Kerckaert J, Montarras D, Pinset C, Lindon C. Overexpressed BCL6 (LAZ3) oncoprotein triggers apoptosis, delays S phase progression and associates with replication foci. Oncogene. 1999;18:5063–5075. - PubMed
-
- Arnold H-H, Winter B. Muscle differentiation: more complexity to the network of myogenic regulators. Curr Opin Genet Dev. 1998;8:539–544. - PubMed
-
- Auradé F, Pinset C, Chafey P, Gros F, Montarras D. Myf5, MyoD, myogenin and MRF4 myogenic derivatives of the embryonic mesenchymal cell line C3H10T1/2 exhibit the same adult muscle phenotype. Differentiation. 1994;55:185–192. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous