

Identification of ubiquilin, a novel presenilin interactor that increases presenilin protein accumulation
- PMID: 11076969
- PMCID: PMC2169435
- DOI: 10.1083/jcb.151.4.847
Identification of ubiquilin, a novel presenilin interactor that increases presenilin protein accumulation
Abstract
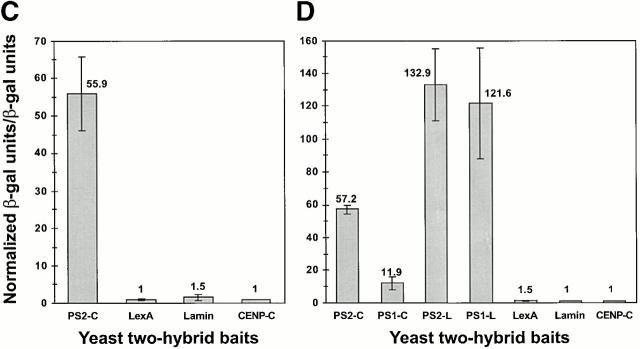
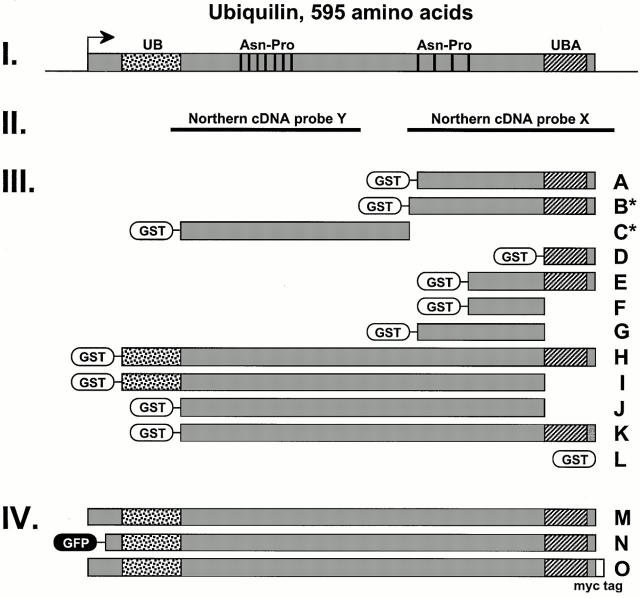
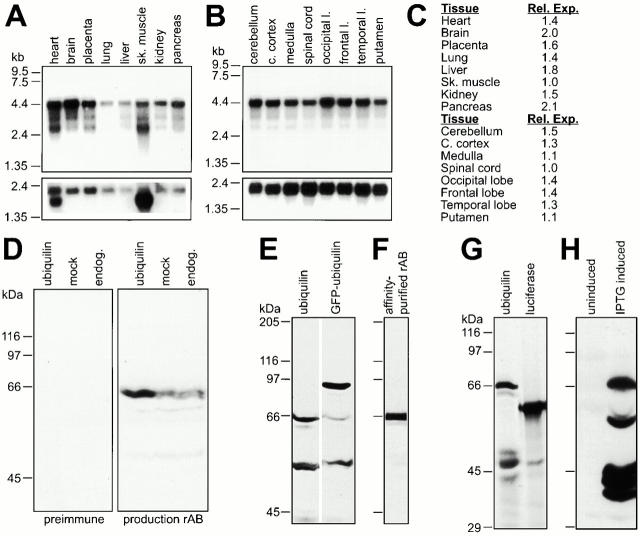
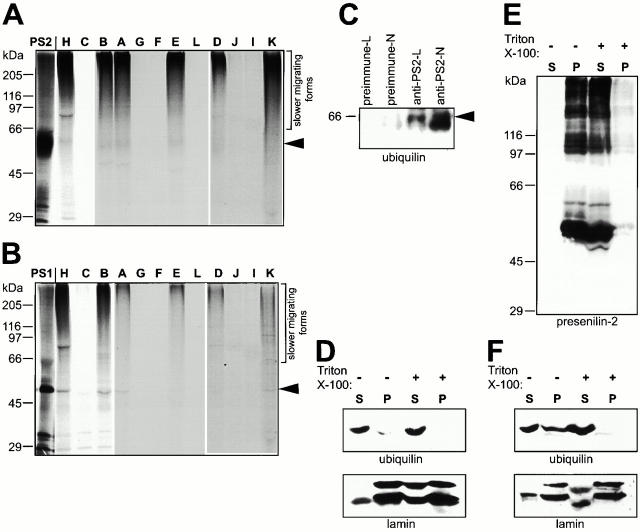
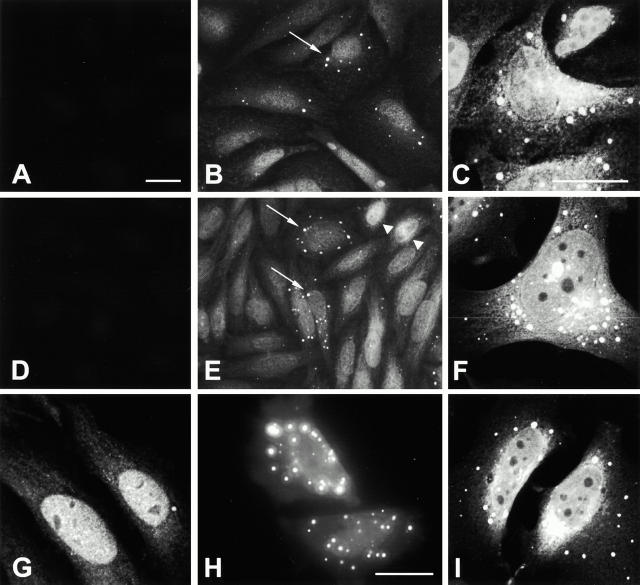
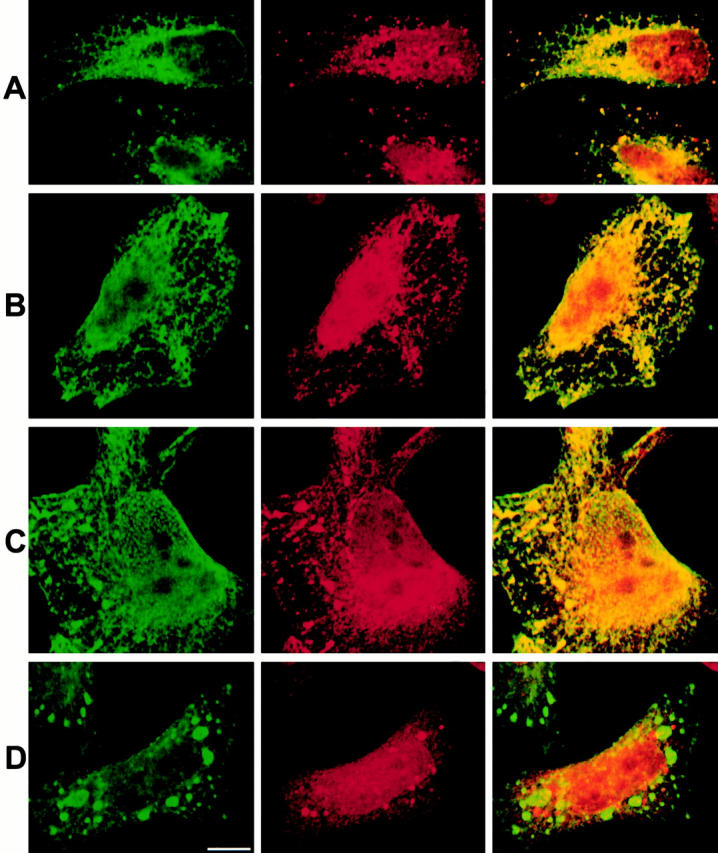
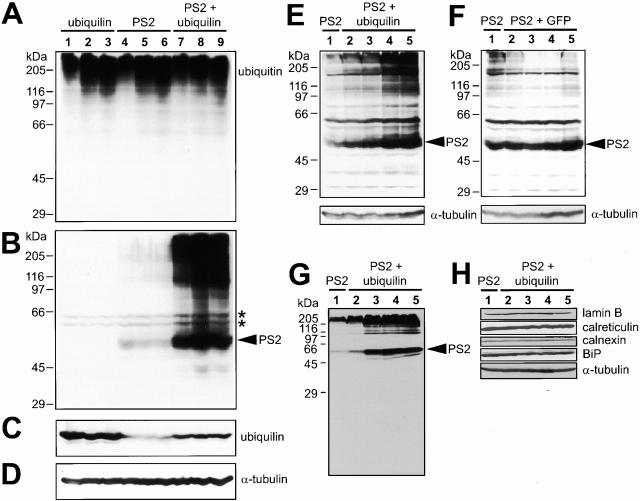
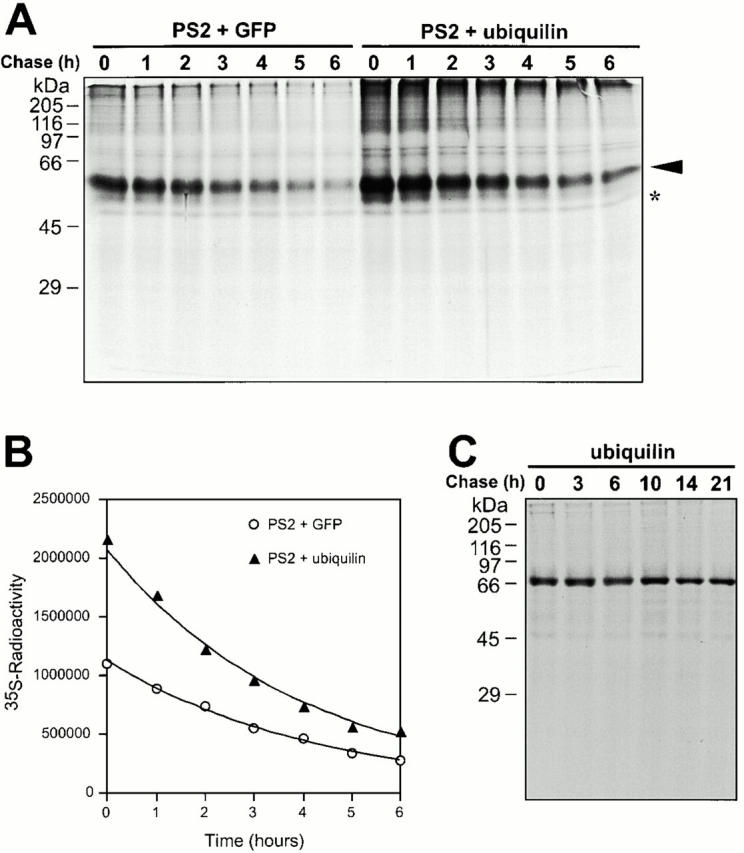
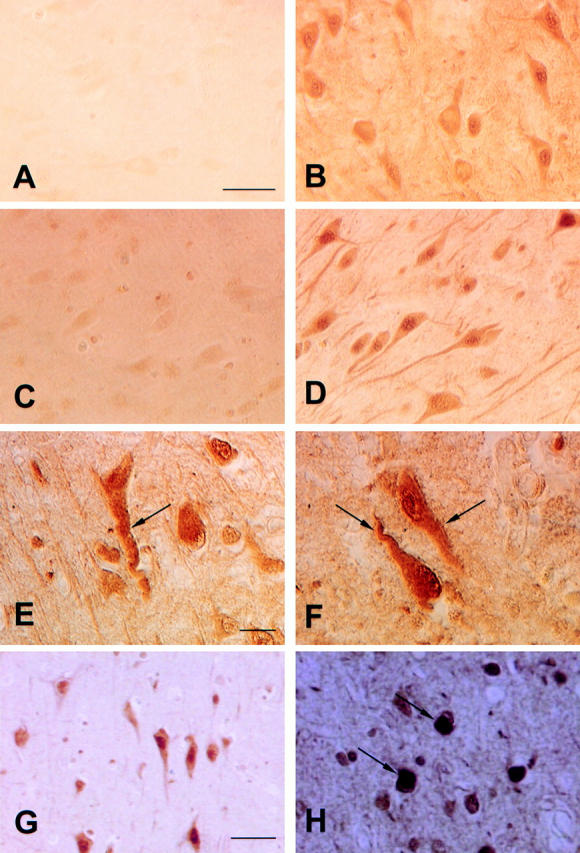
Mutations in the highly homologous presenilin genes encoding presenilin-1 and presenilin-2 (PS1 and PS2) are linked to early-onset Alzheimer's disease (AD). However, apart from a role in early development, neither the normal function of the presenilins nor the mechanisms by which mutant proteins cause AD are well understood. We describe here the properties of a novel human interactor of the presenilins named ubiquilin. Yeast two-hybrid (Y2H) interaction, glutathione S-transferase pull-down experiments, and colocalization of the proteins expressed in vivo, together with coimmunoprecipitation and cell fractionation studies, provide compelling evidence that ubiquilin interacts with both PS1 and PS2. Ubiquilin is noteworthy since it contains multiple ubiquitin-related domains typically thought to be involved in targeting proteins for degradation. However, we show that ubiquilin promotes presenilin protein accumulation. Pulse-labeling experiments indicate that ubiquilin facilitates increased presenilin synthesis without substantially changing presenilin protein half-life. Immunohistochemistry of human brain tissue with ubiquilin-specific antibodies revealed prominent staining of neurons. Moreover, the anti-ubiquilin antibodies robustly stained neurofibrillary tangles and Lewy bodies in AD and Parkinson's disease affected brains, respectively. Our results indicate that ubiquilin may be an important modulator of presenilin protein accumulation and that ubiquilin protein is associated with neuropathological neurofibrillary tangles and Lewy body inclusions in diseased brain.
Figures

References
-
- Braun B.C., Glickman M., Kraft R., Dahlmann B., Kloetzel P.M., Finley D., Schmidt M. The base of the proteasome regulatory particle exhibits chaperone-like activity. Nat. Cell Biol. 1999;1:221–226. - PubMed
-
- Cotman C.W. Apoptosis decision cascades and neuronal degeneration in Alzheimer's disease. Neurobiol. Aging. 1998;19:S29–S32. - PubMed
-
- Davis J.A., Naruse S., Chen H., Eckman C., Younkin S., Price D.L., Borchelt D.R., Sisodia S.S., Wong P.C. An Alzheimer's disease-linked PS1 variant rescues the developmental abnormalities of PS1-deficient embryos. Neuron. 1998;20:603–609. - PubMed
-
- Dieckmann T., Withers-Ward E.S., Jarosinski M.A., Liu C.F., Chen I.S., Feigon J. Structure of a human DNA repair protein UBA domain that interacts with HIV-1 Vpr. Nat. Struct. Biol. 1998;5:1042–1047. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
