

Connexin43 deficiency causes delayed ossification, craniofacial abnormalities, and osteoblast dysfunction
- PMID: 11076975
- PMCID: PMC2169447
- DOI: 10.1083/jcb.151.4.931
Connexin43 deficiency causes delayed ossification, craniofacial abnormalities, and osteoblast dysfunction
Abstract
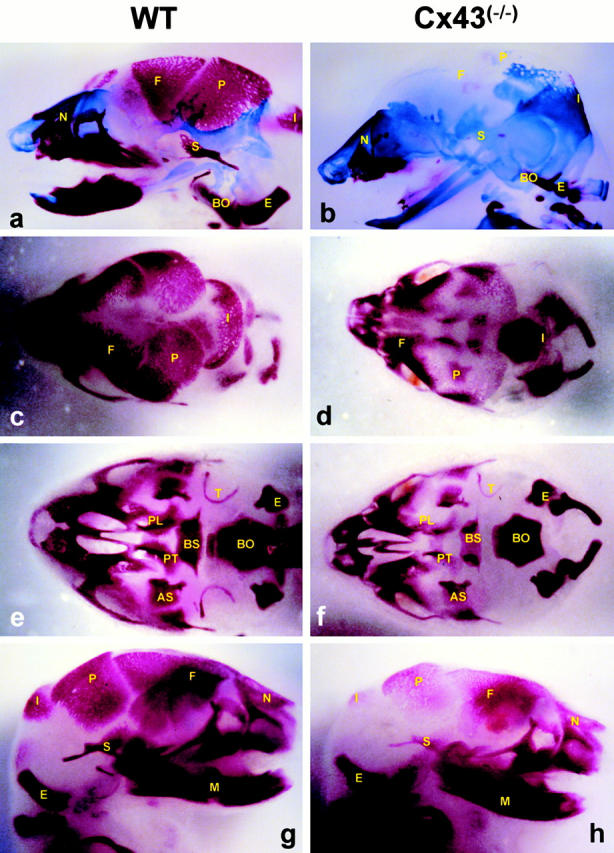
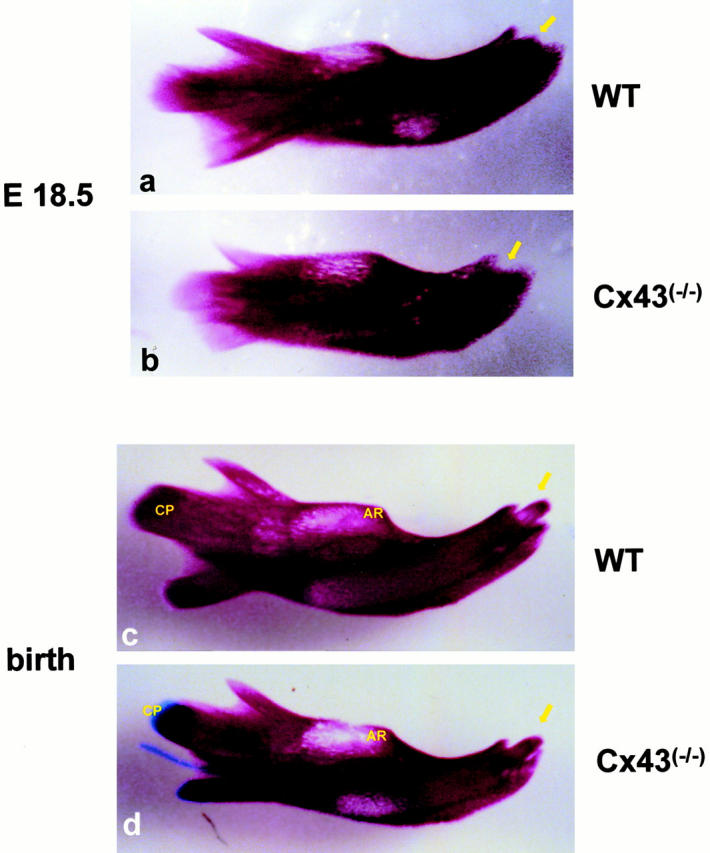
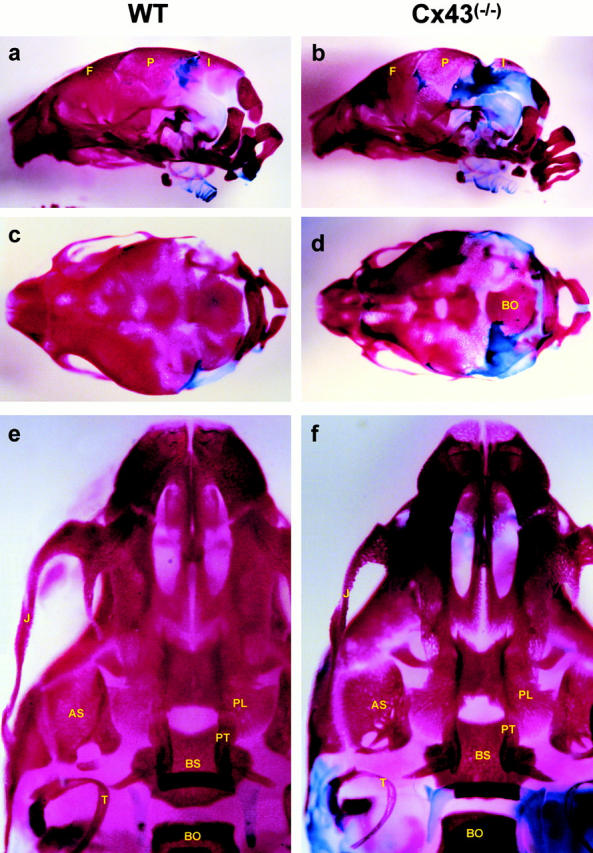
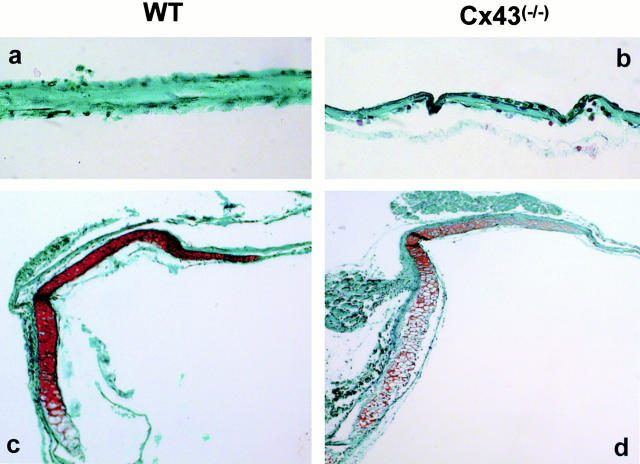
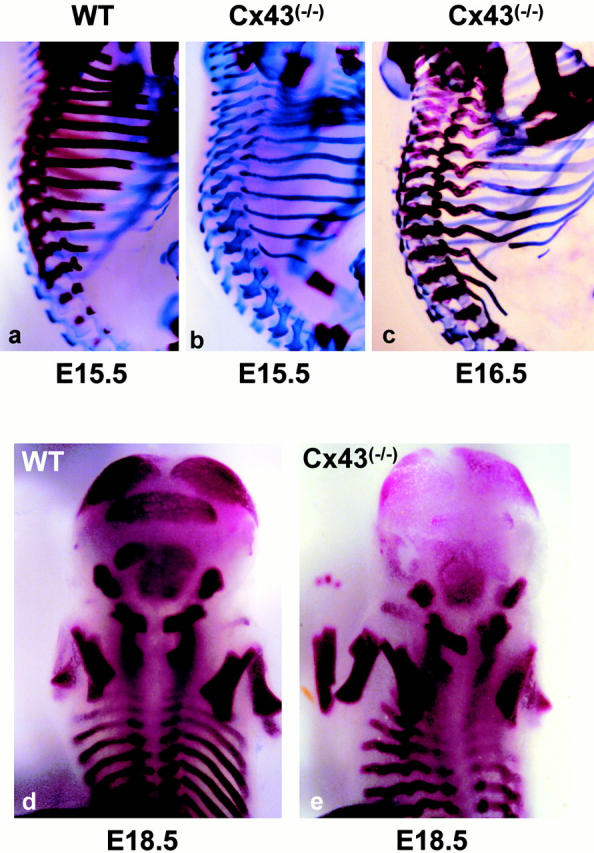
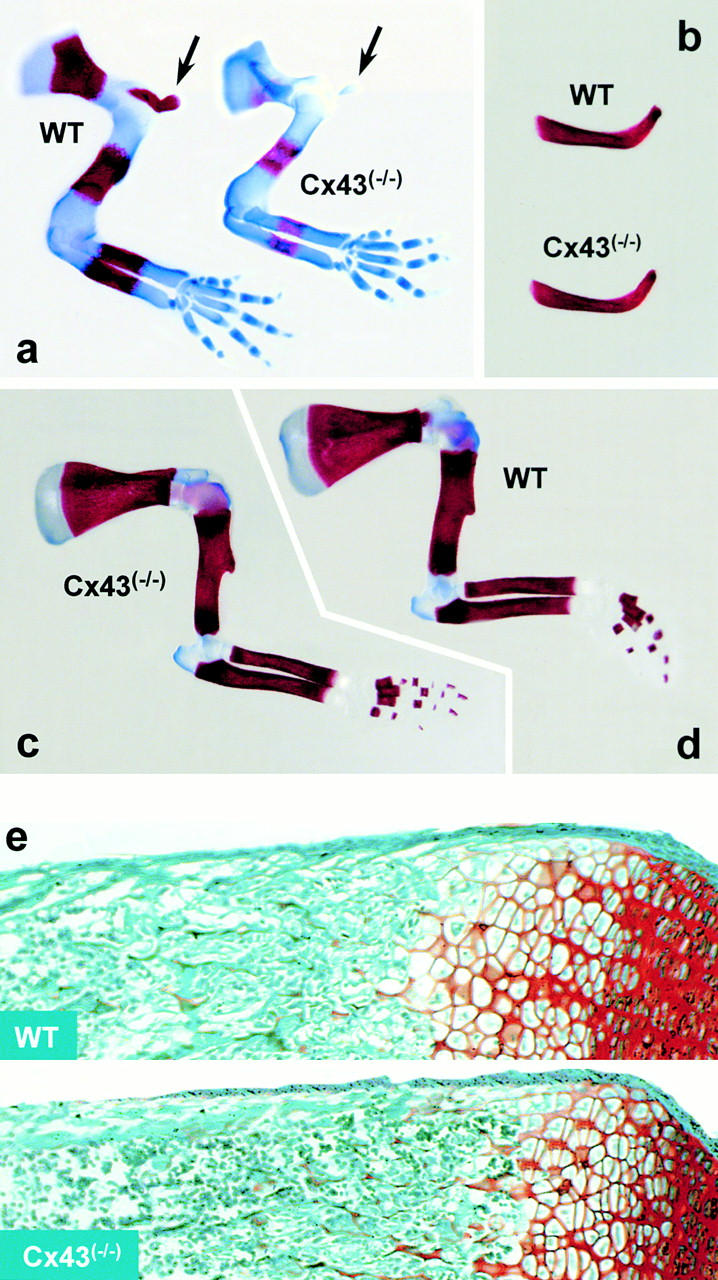
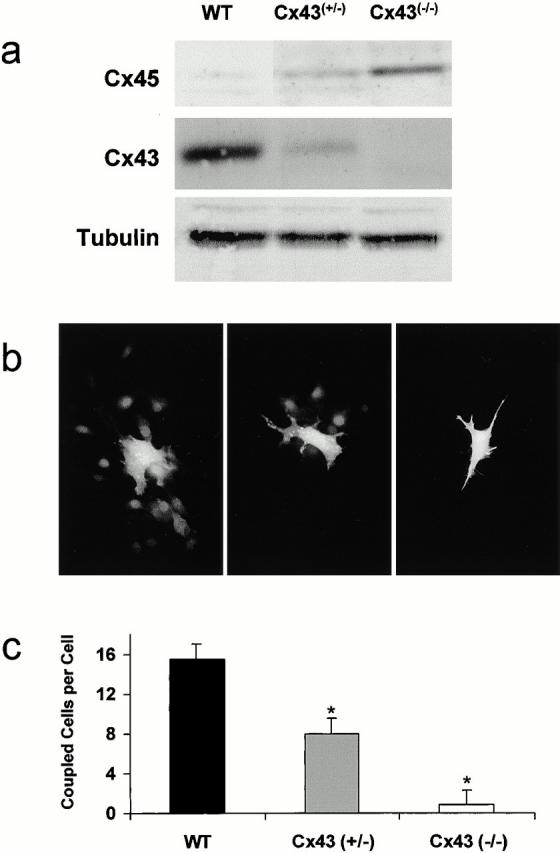
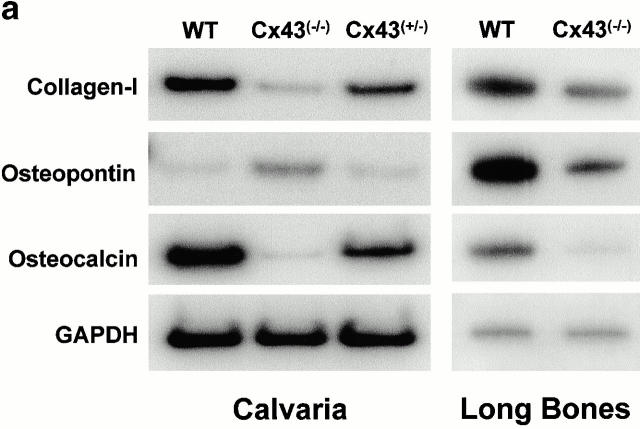
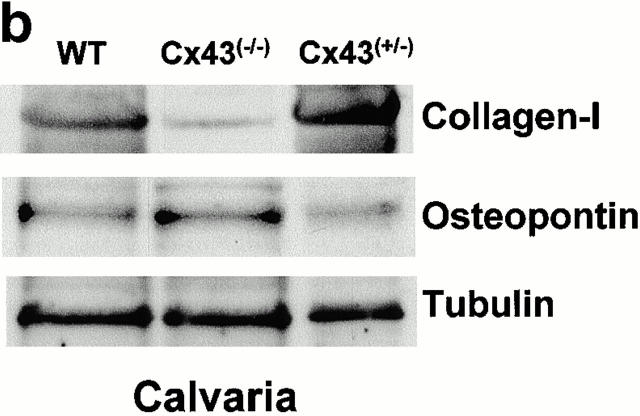
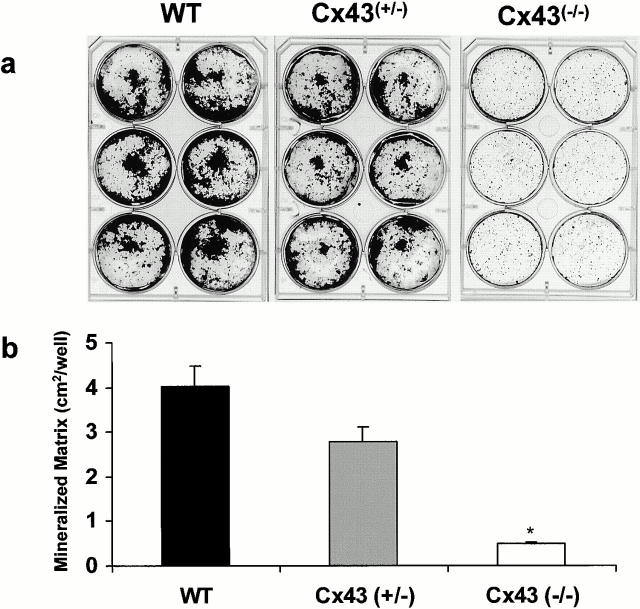
Connexin(Cx)43 is the major gap junction protein present in osteoblasts. We have shown that overexpression of Cx45 in osteoblasts expressing endogenous Cx43 leads to decreased cell-cell communication (Koval, M., S.T. Geist, E.M. Westphale, A.E. Kemendy, R. Civitelli, E.C. Beyer, and T.H. Steinberg. 1995. J. Cell Biol. 130:987-995) and transcriptional downregulation of several osteoblastic differentiation markers (Lecanda, F., D.A. Towler, K. Ziambaras, S.-L. Cheng, M. Koval, T.H. Steinberg, and R. Civitelli. 1998. Mol. Biol. Cell 9:2249-2258). Here, using the Cx43-null mouse model, we determined whether genetic deficiency of Cx43 affects skeletal development in vivo. Both intramembranous and endochondral ossification of the cranial vault were delayed in the mutant embryos, and cranial bones originating from migratory neural crest cells were also hypoplastic, leaving an open foramen at birth. Cx43-deficient animals also exhibited retarded ossification of the clavicles, ribs, vertebrae, and limbs, demonstrating that skeletal abnormalities are not restricted to a neural crest defect. However, the axial and appendicular skeleton of Cx43-null animals were essentially normal at birth. Cell to cell diffusion of calcein was poor among Cx43-deficient osteoblasts, whose differentiated phenotypic profile and mineralization potential were greatly impaired, compared with wild-type cells. Therefore, in addition to the reported neural crest cell defect, lack of Cx43 also causes a generalized osteoblast dysfunction, leading to delayed mineralization and skull abnormalities. Cell to cell signaling, mediated by Cx43 gap junctions, was critical for normal osteogenesis, craniofacial development, and osteoblastic function.
Figures
References
-
- Becker D.L., McGonnell I., Makarenkova H.P., Patel K., Tickle C., Lorimer J., Green C.R. Roles for alpha 1 connexin in morphogenesis of chick embryos revealed using a novel antisense approach. Dev. Genet. 1999;24:33–42. - PubMed
-
- Bradford M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein dye binding. Anal. Biochem. 1976;72:248–254. - PubMed
-
- Chen H., Ovchinnikov D., Pressman C.L., Aulehla A., Lun Y., Johnson R.L. Multiple calvarial defects in lmx1b mutant mice. Dev. Genet. 1998;22:314–320. - PubMed
-
- Chen Y., Bei M., Woo I., Satokata I., Maas R. Msx1 controls inductive signaling in mammalian tooth morphogenesis. Development. 1996;122:3035–3044. - PubMed
-
- Cheng S.-L., Yang J.W., Rifas L., Zhang S.-F., Avioli L.V. Differentiation of human bone marrow osteogenic stromal cells in vitroinduction of the osteoblast phenotype by dexamethasone. Endocrinology. 1994;134:277–286. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
