

Protein folding in the periplasm in the absence of primary oxidant DsbA: modulation of redox potential in periplasmic space via OmpL porin
- PMID: 11080145
- PMCID: PMC305838
- DOI: 10.1093/emboj/19.22.5980
Protein folding in the periplasm in the absence of primary oxidant DsbA: modulation of redox potential in periplasmic space via OmpL porin
Erratum in
- EMBO J. 2004 Oct 1;23(19):3907
Abstract
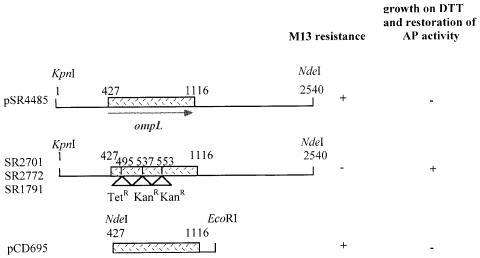
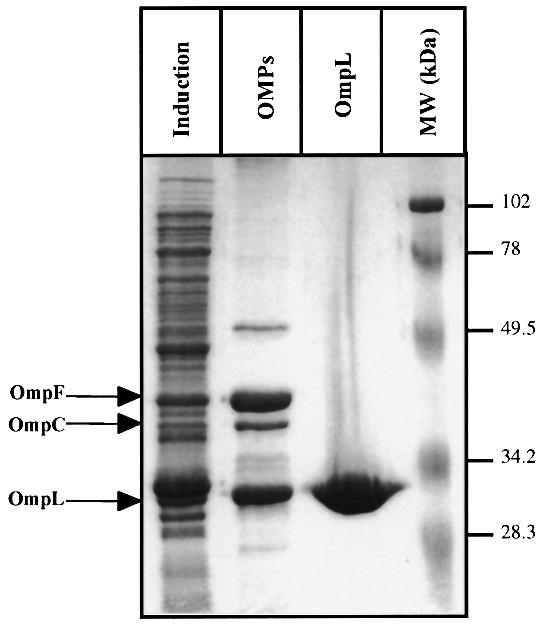
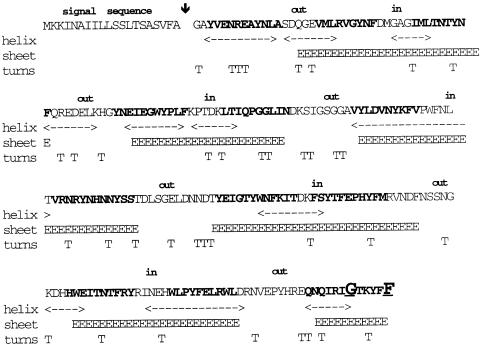
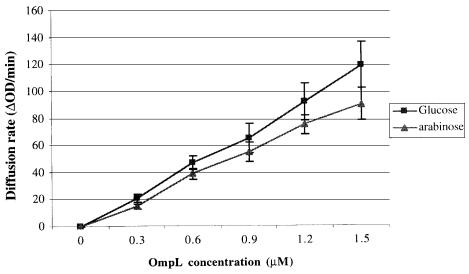
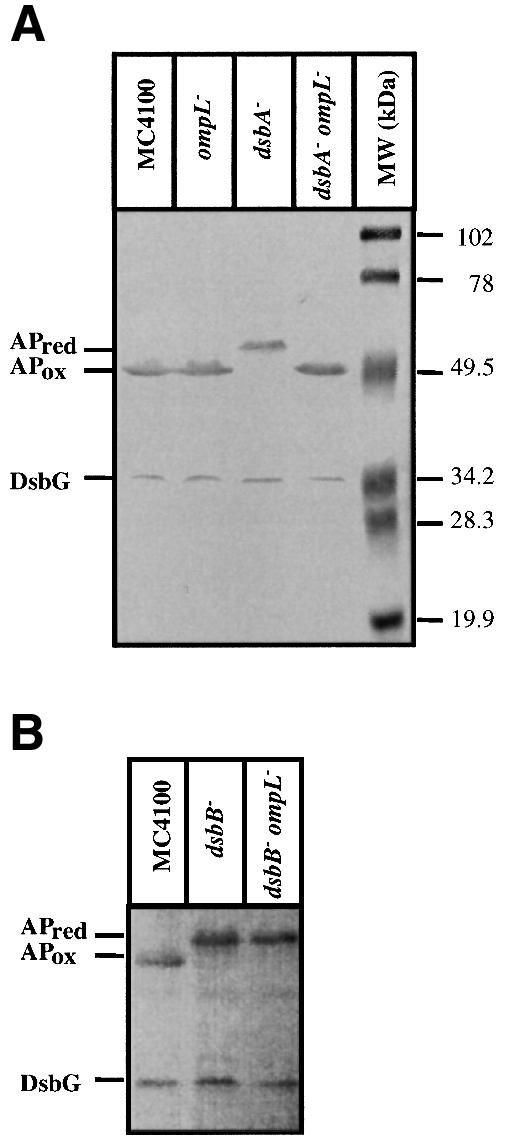
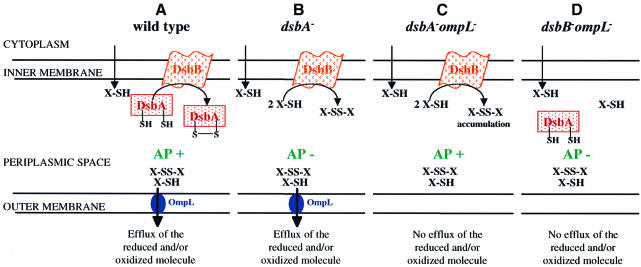
Disulfide bond formation in Escherichia coli is a catalyzed reaction accomplished by DsbA. We found that null mutations in a new porin gene, ompL, allowed a total bypass of the DsbA requirement for protein oxidation. These mutations acted as extragenic null suppressors for dsbA, and restored normal folding of alkaline phosphatase and relieved sensitivity to dithiothreitol. ompL dsbA double mutants were completely like wild-type mutants in terms of motility and lack of mucoidy. This suppression was not dependent on DsbC and DsbG, since the oxidation status of these proteins was unaltered in ompL dsbA strains. Purified OmpL allowed diffusion of small solutes, including sugars, but the suppression was not dependent on the carbon sources used. Suppression by ompL null mutations required DsbB, leading us to propose a hypothesis that DsbB oxidizes yet unidentified, low-molecular-weight redox agents in the periplasm. These oxidized agents accumulate and substitute for DsbA if their leakage into the medium is prevented by the absence of OmpL, presumed to form a specific channel for their diffusion.
Figures
Similar articles
-
The OmpL porin does not modulate redox potential in the periplasmic space of Escherichia coli.EMBO J. 2003 Apr 1;22(7):1461-6. doi: 10.1093/emboj/cdg152. EMBO J. 2003. PMID: 12660153 Free PMC article.
-
A new Escherichia coli gene, dsbG, encodes a periplasmic protein involved in disulphide bond formation, required for recycling DsbA/DsbB and DsbC redox proteins.Mol Microbiol. 1997 Oct;26(1):121-32. doi: 10.1046/j.1365-2958.1997.5581925.x. Mol Microbiol. 1997. PMID: 9383195
-
Complementation of DsbA deficiency with secreted thioredoxin variants reveals the crucial role of an efficient dithiol oxidant for catalyzed protein folding in the bacterial periplasm.EMBO J. 1999 Jun 15;18(12):3271-81. doi: 10.1093/emboj/18.12.3271. EMBO J. 1999. PMID: 10369668 Free PMC article.
-
[Escherichia coli disulfide-forming related proteins: structures, functions and their application in gene engineering for expressing heterologous proteins in Escherichia coli].Sheng Wu Gong Cheng Xue Bao. 2002 May;18(3):261-6. Sheng Wu Gong Cheng Xue Bao. 2002. PMID: 12192853 Review. Chinese.
-
Periplasmic chaperones used to enhance functional secretion of proteins in E. coli.Methods Mol Biol. 2011;705:211-24. doi: 10.1007/978-1-61737-967-3_12. Methods Mol Biol. 2011. PMID: 21125388 Review.
Cited by
-
Molecular basis of bacterial outer membrane permeability revisited.Microbiol Mol Biol Rev. 2003 Dec;67(4):593-656. doi: 10.1128/MMBR.67.4.593-656.2003. Microbiol Mol Biol Rev. 2003. PMID: 14665678 Free PMC article. Review.
-
Stress adaptation under in vitro evolution influences survival and metabolic phenotypes of clinical and environmental strains of Vibrio cholerae El-Tor.Microbiol Spectr. 2025 Mar 4;13(3):e0121124. doi: 10.1128/spectrum.01211-24. Epub 2025 Feb 11. Microbiol Spectr. 2025. PMID: 39932327 Free PMC article.
-
Salmonella produces an O-antigen capsule regulated by AgfD and important for environmental persistence.J Bacteriol. 2006 Nov;188(22):7722-30. doi: 10.1128/JB.00809-06. J Bacteriol. 2006. PMID: 17079680 Free PMC article.
-
The Whole Is Bigger than the Sum of Its Parts: Drug Transport in the Context of Two Membranes with Active Efflux.Chem Rev. 2021 May 12;121(9):5597-5631. doi: 10.1021/acs.chemrev.0c01137. Epub 2021 Feb 17. Chem Rev. 2021. PMID: 33596653 Free PMC article.
-
Function and expression of an N-acetylneuraminic acid-inducible outer membrane channel in Escherichia coli.J Bacteriol. 2005 Mar;187(6):1959-65. doi: 10.1128/JB.187.6.1959-1965.2005. J Bacteriol. 2005. PMID: 15743943 Free PMC article.
References
-
- Andersen C., Matthey-Dupraz,A., Missiakas,D. and Raina,S. (1997) A new Escherichia coli gene, dsbG, encodes a periplasmic protein involved in disulphide bond formation, required for recycling DsbA/DsbB and DsbC redox proteins. Mol. Microbiol., 26, 121–132. - PubMed
-
- Bader M., Muse,W., Ballou,D.P., Gassner,C. and Bardwell,J.C. (1999) Oxidative protein folding is driven by the electron transport system. Cell, 98, 217–227. - PubMed
-
- Bardwell J.C.A., McGovern,K. and Beckwith,J. (1991) Identification of a protein required for disulfide bond formation in vivo. Cell, 67, 581–589. - PubMed
-
- Bessette P., Cotto,J., Gilbert,H. and Georgiou,G. (1999) In vivo and in vitro function of the Escherichia coli periplasmic cysteine oxidoreductase DsbG. J. Biol. Chem., 274, 7784–7792. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
