

Pontin52 and reptin52 function as antagonistic regulators of beta-catenin signalling activity
- PMID: 11080158
- PMCID: PMC305835
- DOI: 10.1093/emboj/19.22.6121
Pontin52 and reptin52 function as antagonistic regulators of beta-catenin signalling activity
Abstract
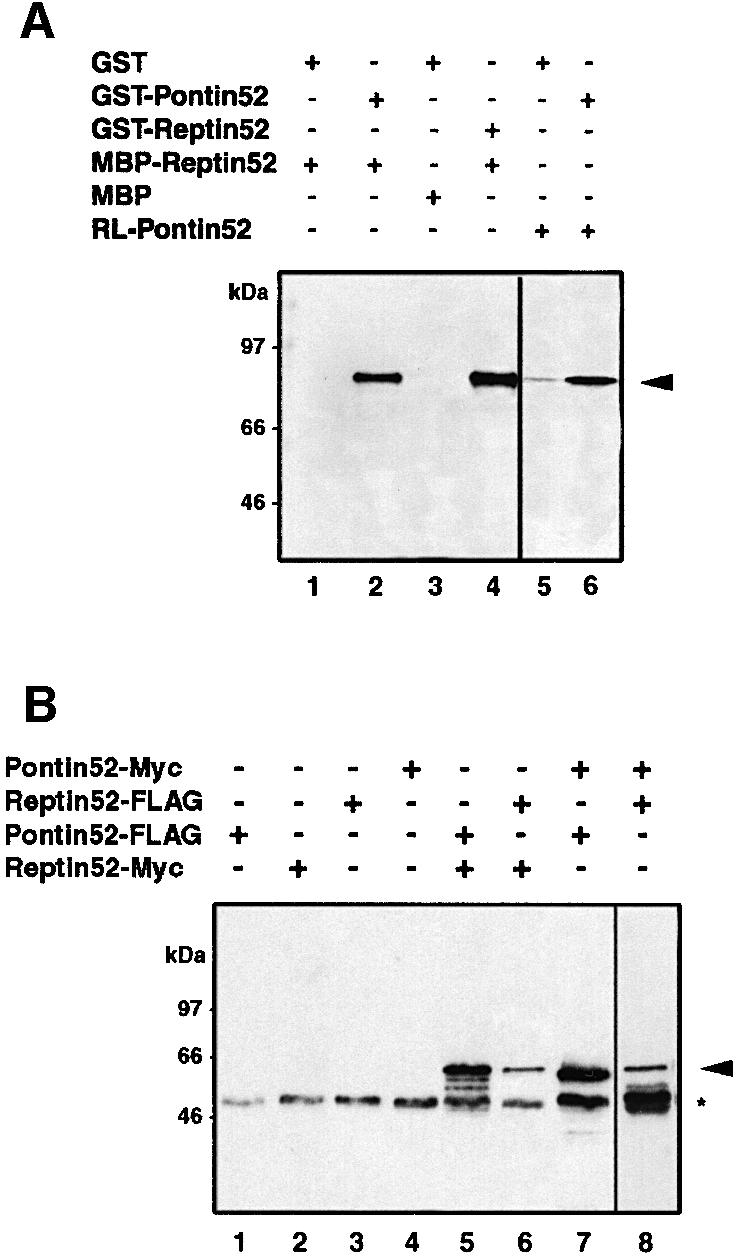
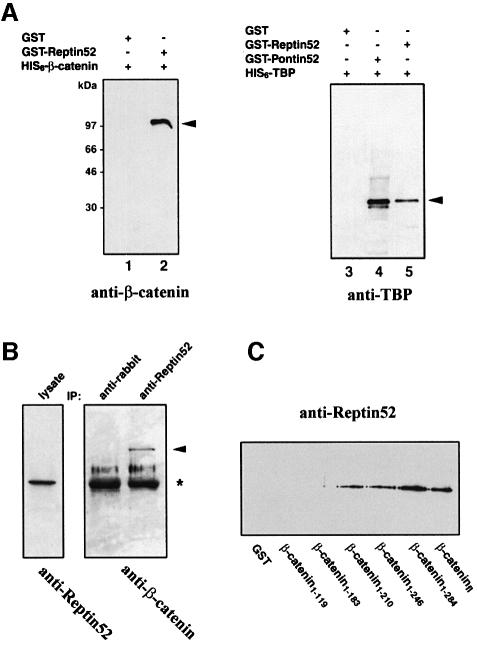
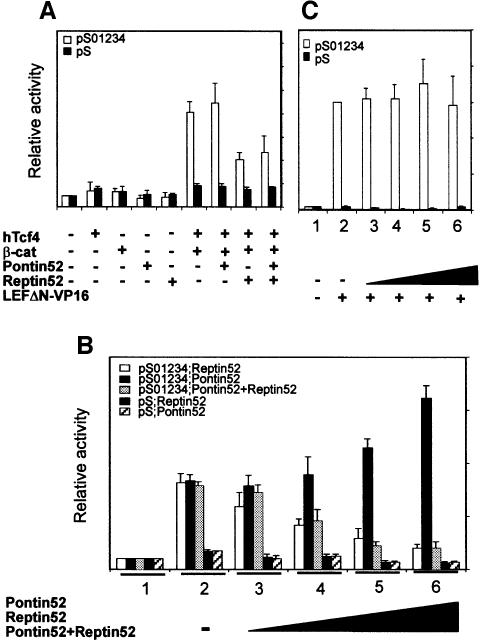
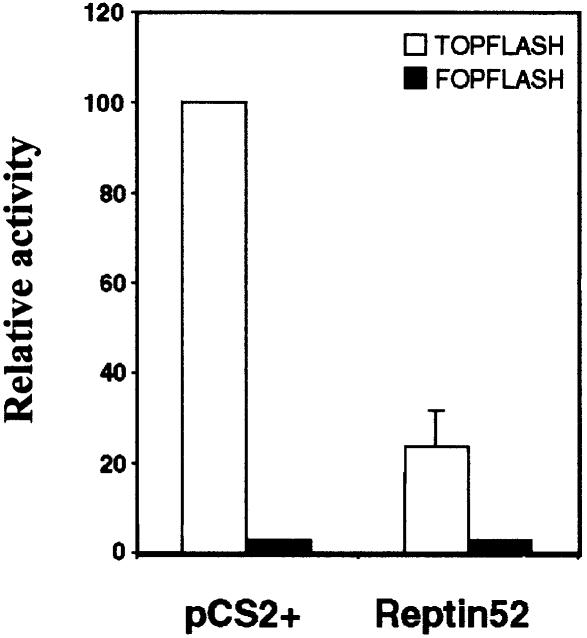
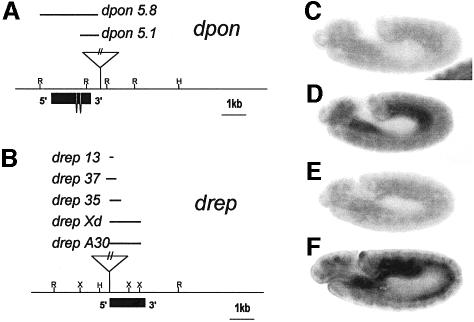
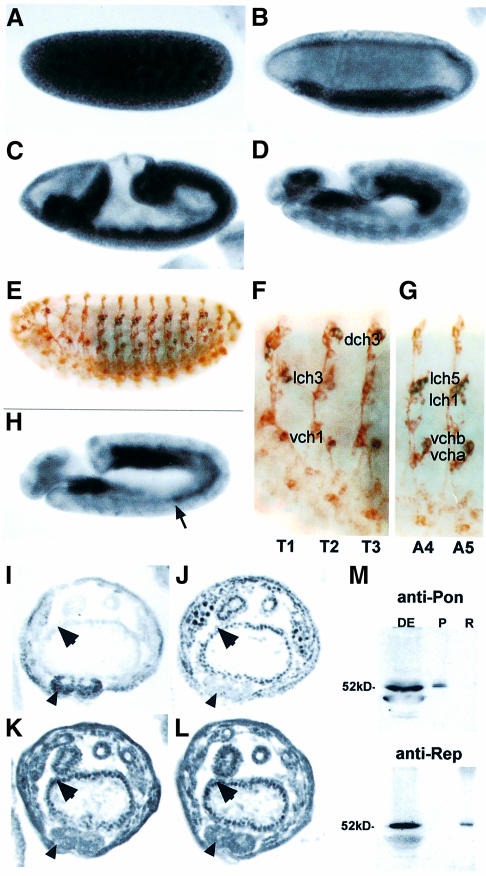
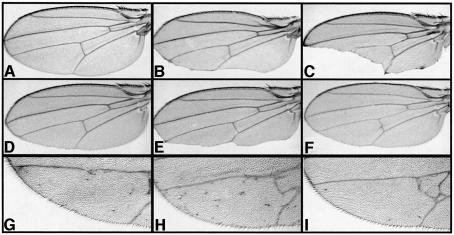
In Wnt-stimulated cells, beta-catenin becomes stabilized in the cytoplasm, enters the nucleus and interacts with HMG box transcription factors of the lymphoid-enhancing factor-1 (LEF-1)/T-cell factor (TCF) family, thereby stimulating the transcription of specific target genes. We recently identified Pontin52 as a nuclear protein interacting with beta-catenin and the TATA-box binding protein (TBP), suggesting its involvement in regulating beta-catenin-mediated transactivation. Here, we report the identification of Reptin52 as an interacting partner of Pontin52. Highly homologous to Pontin52, Reptin52 likewise binds beta-catenin and TBP. Using reporter gene assays, we show that the two proteins antagonistically influence the transactivation potential of the beta-catenin-TCF complex. Furthermore, we demonstrate the evolutionary conservation of this mechanism in Drosophila. dpontin and dreptin are essential genes that act antagonistically in the control of Wingless signalling in vivo. These results indicate that the opposite action of Pontin52 and Reptin52 on beta-catenin-mediated transactivation constitutes an additional mechanism for the control of the canonical Wingless/Wnt pathway.
Figures

References
-
- Akimaru H., Hou,D.X. and Ishii,S. (1997) Drosophila CBP is required for dorsal-dependent twist gene expression. Nature Genet., 17, 211–214. - PubMed
-
- Alexandre E., Graba,Y., Fasano,L., Gallet,A., Perrin,L., De Zulueta,P., Pradel,J., Kerridge,S. and Jacq,B. (1996) The Drosophila Teashirt homeotic protein is a DNA-binding protein and modulo, a HOM-C regulated modifier of variegation, is a likely candidate for being a direct target gene. Mech. Dev., 59, 191–204. - PubMed
-
- Bhanot P., Fish,M., Jemison,J.A., Nusse,R., Nathans,J. and Cadigan,K.M. (1999) Frizzled and Dfrizzled-2 function as redundant receptors for Wingless during Drosophila embryonic development. Development, 126, 4175–4186. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
