

The machinery of local Ca2+ signalling between sarco-endoplasmic reticulum and mitochondria
- PMID: 11080252
- PMCID: PMC2270182
- DOI: 10.1111/j.1469-7793.2000.00069.x
The machinery of local Ca2+ signalling between sarco-endoplasmic reticulum and mitochondria
Abstract
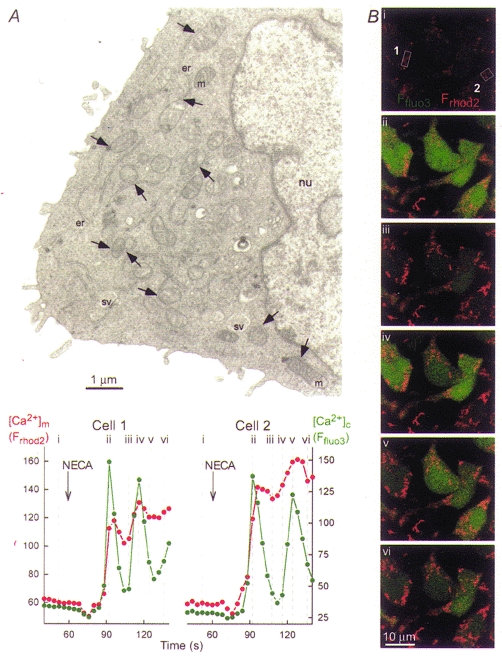
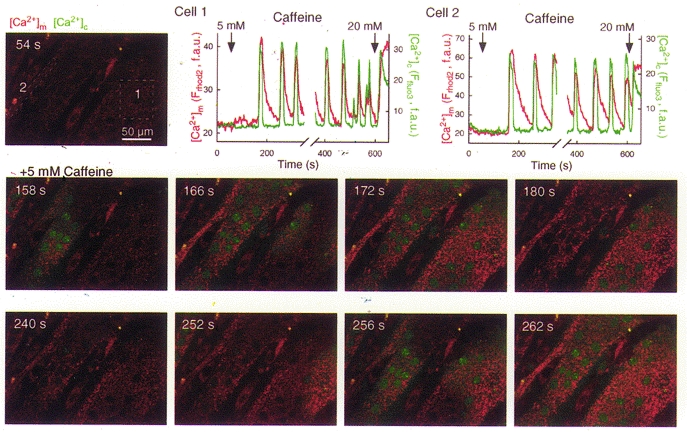
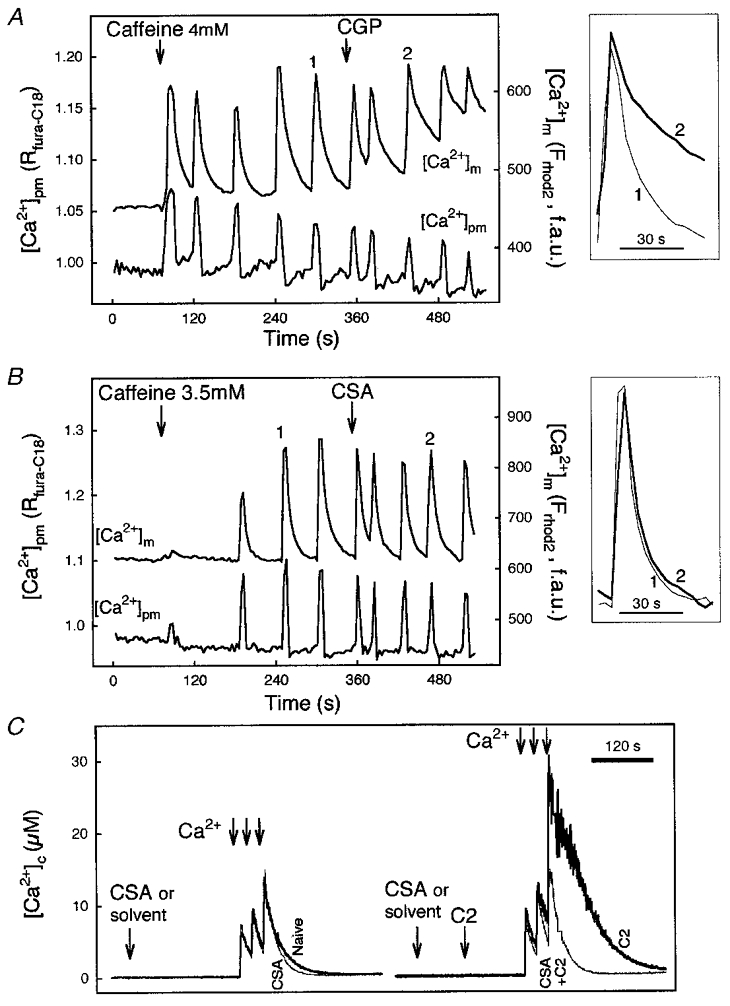
Growing evidence suggests that propagation of cytosolic [Ca2+] ([Ca2+]c) spikes and oscillations to the mitochondria is important for the control of fundamental cellular functions. Delivery of [Ca2+]c spikes to the mitochondria may utilize activation of the mitochondrial Ca2+ uptake sites by the large local [Ca2+]c rise occurring in the vicinity of activated sarco-endoplasmic reticulum (SR/ER) Ca2+ release channels. Although direct measurement of the local [Ca2+]c sensed by the mitochondria has been difficult, recent studies shed some light onto the molecular mechanism of local Ca2+ communication between SR/ER and mitochondria. Subdomains of the SR/ER are in close contact with mitochondria and display a concentration of Ca2+ release sites, providing the conditions for an effective delivery of released Ca2+ to the mitochondrial targets. Furthermore, many functional properties of the signalling between SR/ER Ca2+ release sites and mitochondrial Ca2+ uptake sites, including transient microdomains of high [Ca2+], saturation of mitochondrial Ca2+ uptake sites by released Ca2+, connection of multiple release sites to each uptake site and quantal transmission, are analogous to the features of the coupling between neurotransmitter release sites and postsynaptic receptors in synaptic transmission. As such, Ca2+ signal transmission between SR/ER and mitochondria may utilize discrete communication sites and a closely related functional architecture to that used for synaptic signal propagation between cells.
Figures
References
-
- Altschuld RA, Hohl CM, Castillo LC, Garleb AA, Starling RC, Brierley GP. Cyclosporin inhibits mitochondrial calcium efflux in isolated adult rat ventricular cardiomyocytes. American Journal of Physiology. 1992;262:1699–1704. - PubMed
-
- Babcock DF, Hille B. Mitochondrial oversight of cellular Ca2+ signalling. Current Opinions in Neurobiology. 1998;8:398–404. - PubMed
-
- Bereiter-Hahn J, Voth M. Dynamics of mitochondria in living cells: shape changes, dislocations, fusion, and fission of mitochondria. Microscopy Research and Technique. 1994;27:198–219. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous