

Cryo-scanning electron microscopy observations of vessel content during transpiration in walnut petioles. Facts or artifacts?
- PMID: 11080296
- PMCID: PMC59218
- DOI: 10.1104/pp.124.3.1191
Cryo-scanning electron microscopy observations of vessel content during transpiration in walnut petioles. Facts or artifacts?
Abstract
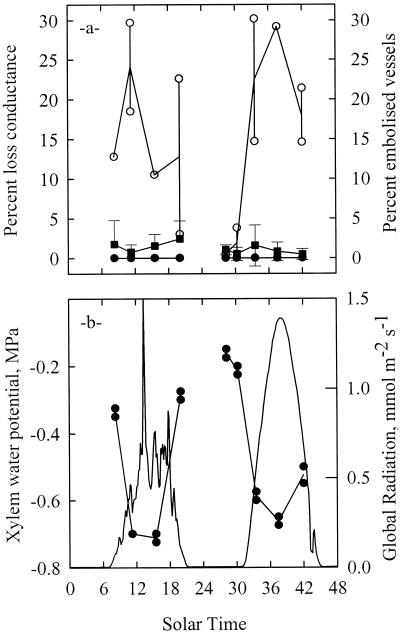
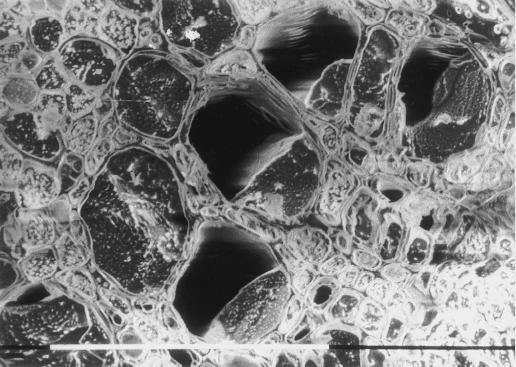
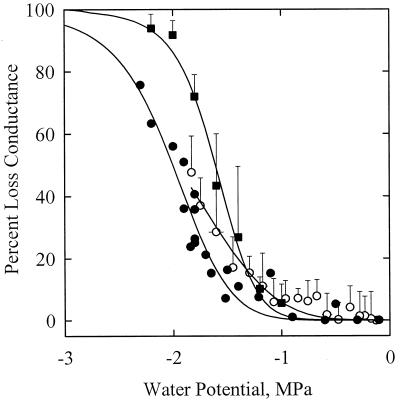
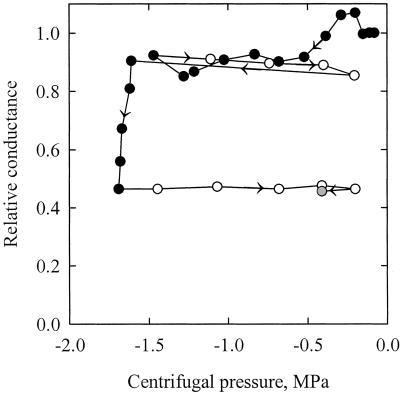
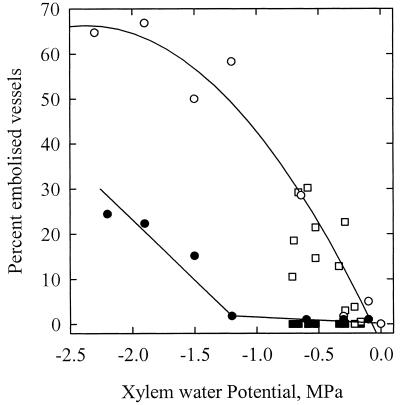
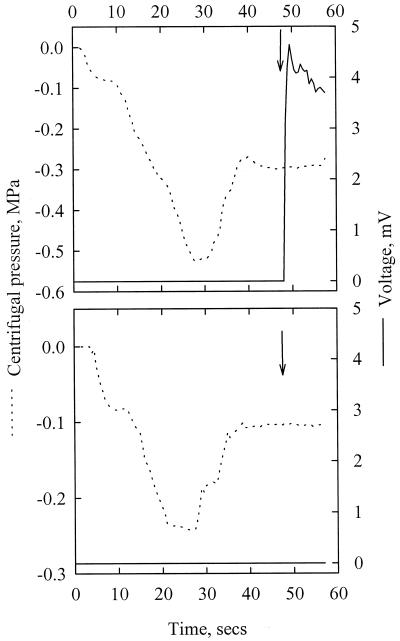
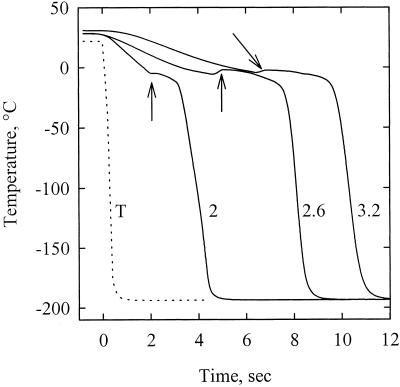
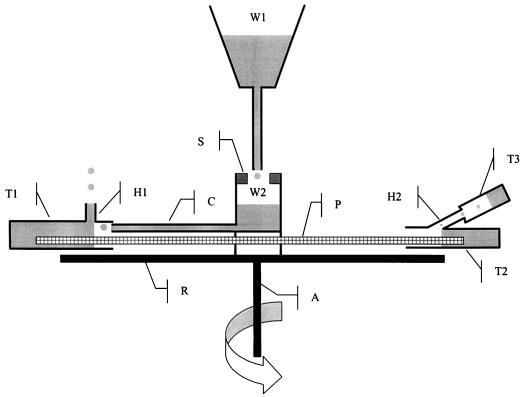
The current controversy about the "cohesion-tension" of water ascent in plants arises from the recent cryo-scanning electron microscopy (cryo-SEM) observations of xylem vessels content by Canny and coworkers (1995). On the basis of these observations it has been claimed that vessels were emptying and refilling during active transpiration in direct contradiction to the previous theory. In this study we compared the cryo-SEM data with the standard hydraulic approach on walnut (Juglans regia) petioles. The results of the two techniques were in clear conflict and could not both be right. Cryo-SEM observations of walnut petioles frozen intact on the tree in a bath of liquid nitrogen (LN(2)) suggested that vessel cavitation was occurring and reversing itself on a diurnal basis. Up to 30% of the vessels were embolized at midday. In contrast, the percentage of loss of hydraulic conductance (PLC) of excised petiole segments remained close to 0% throughout the day. To find out which technique was erroneous we first analyzed the possibility that PLC values were rapidly returned to zero when the xylem pressures were released. We used the centrifugal force to measure the xylem conductance of petiole segments exposed to very negative pressures and established the relevance of this technique. We then analyzed the possibility that vessels were becoming partially air-filled when exposed to LN(2). Cryo-SEM observations of petiole segments frozen shortly after their xylem pressure was returned to atmospheric values agreed entirely with the PLC values. We confirmed, with water-filled capillary tubes exposed to a large centrifugal force, that it was not possible to freeze intact their content with LN(2). We concluded that partially air-filled conduits were artifacts of the cryo-SEM technique in our study. We believe that the cryo-SEM observations published recently should probably be reconsidered in the light of our results before they may be used as arguments against the cohesion-tension theory.
Figures
References
-
- Alder NN, Pockman WT, Sperry JS, Nuismer S. Use of centrifugal force in the study of xylem cavitation. J Exp Bot. 1997;48:665–674.
-
- Berndt ML, McCully ME, Canny MJ. Is xylem embolism and refilling involved in the rapid wilting and recovery of plants following root cooling and rewarming? Plant Biol. 1999;1:506–515.
-
- Böhm J. Capillarität und saftsteigen. Ber Dtsch Bot Ges. 1893;11:203–212.
-
- Briggs LI. Limiting negative pressure of water. J Appl Phys. 1950;21:721–722.
-
- Buchard C, McCully ME, Canny MJ. Daily embolism and refilling of root xylem vessels in three dicotyledonous crop plants. Agronomie. 1999;19:97–106.
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
