

Permeability and channel-mediated transport of boric acid across membrane vesicles isolated from squash roots
- PMID: 11080310
- PMCID: PMC59232
- DOI: 10.1104/pp.124.3.1349
Permeability and channel-mediated transport of boric acid across membrane vesicles isolated from squash roots
Abstract
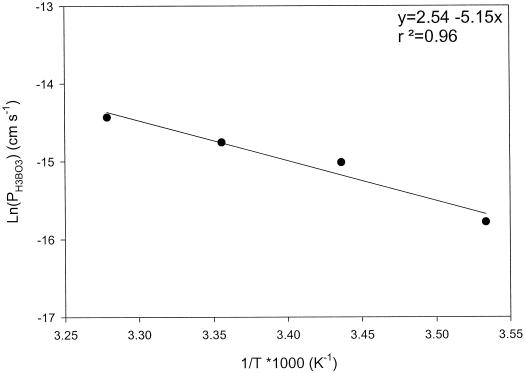
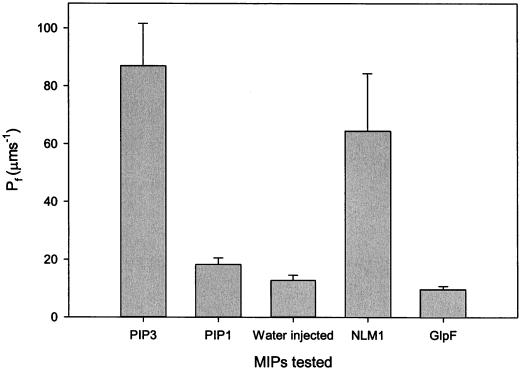
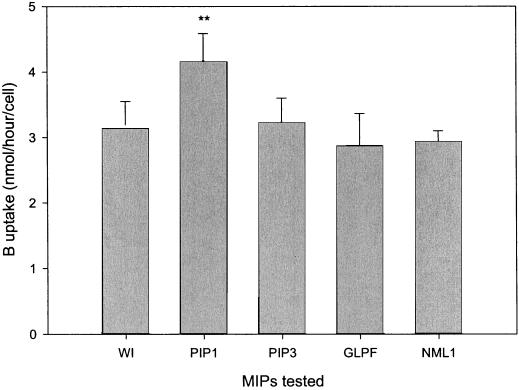
Boron is an essential micronutrient for plant growth and the boron content of plants differs greatly, but the mechanism(s) of its uptake into cells is not known. Boron is present in the soil solution as boric acid and it is in this form that it enters the roots. We determined the boron permeability coefficient of purified plasma membrane vesicles obtained from squash (Cucurbita pepo) roots and found it to be 3 x 10(-7) +/-1.4 x 10(-8) cm s(-1), six times higher than the permeability of microsomal vesicles. Boric acid permeation of the plasma membrane vesicles was partially inhibited (30%-39%) by mercuric chloride and phloretin, a non-specific channel blocker. The inhibition by mercuric chloride was readily reversible by 2-mercaptoethanol. The energy of activation for boron transport into the plasma membrane vesicles was 10.2 kcal mol(-1). Together these data indicate that boron enters plant cells in part by passive diffusion through the lipid bilayer of the plasma membrane and in part through proteinaceous channels. Expression of the major intrinsic protein (MIP) PIP1 in Xenopus laevis oocytes resulted in a 30% increase in the boron permeability of the oocytes. Other MIPs tested (PIP3, MLM1, and GlpF) did not have this effect. We postulate that certain MIPs, like those that have recently been shown to transport small neutral solutes, may also be the channels through which boron enters plant cells.
Figures
References
-
- Agre P, Bonhivers M, Borgnia MJ. The aquaporins, blueprints for cellular plumbing systems. J Biol Chem. 1998;273:14659–14662. - PubMed
-
- Agre P, Mathai JC, Smith BL, Preston GM. Functional analyses of aquaporin water channel proteins. Methods Enzymol. 1999;294:550–572. - PubMed
-
- Barone LM, Shih C, Wasserman BP. Mercury-induced conformational changes and identification of conserved surface loops in plasma membrane aquaporins from higher plants: topology of PMIP31 from Beta vulgaris L. J Biol Chem. 1997;272:30672–30677. - PubMed
-
- Biela A, Grote K, Otto B, Hoth S, Hedrich R, Kaldenhoff R. The Nicotiana tabacum plasma membrane aquaporin NtAQP1 is mercury-insensitive and permeable for glycerol. Plant J. 1999;18:565–570. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
