

Dual role of signaling pathways leading to Ca(2+) and cyclic AMP elevation in host cell invasion by Trypanosoma cruzi
- PMID: 11083771
- PMCID: PMC97756
- DOI: 10.1128/IAI.68.12.6602-6610.2000
Dual role of signaling pathways leading to Ca(2+) and cyclic AMP elevation in host cell invasion by Trypanosoma cruzi
Abstract
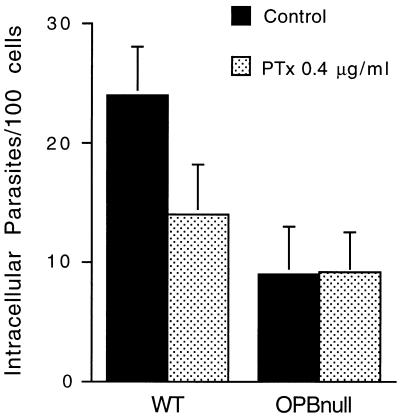
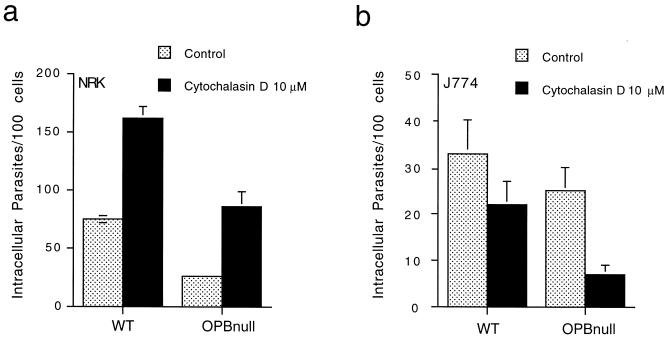
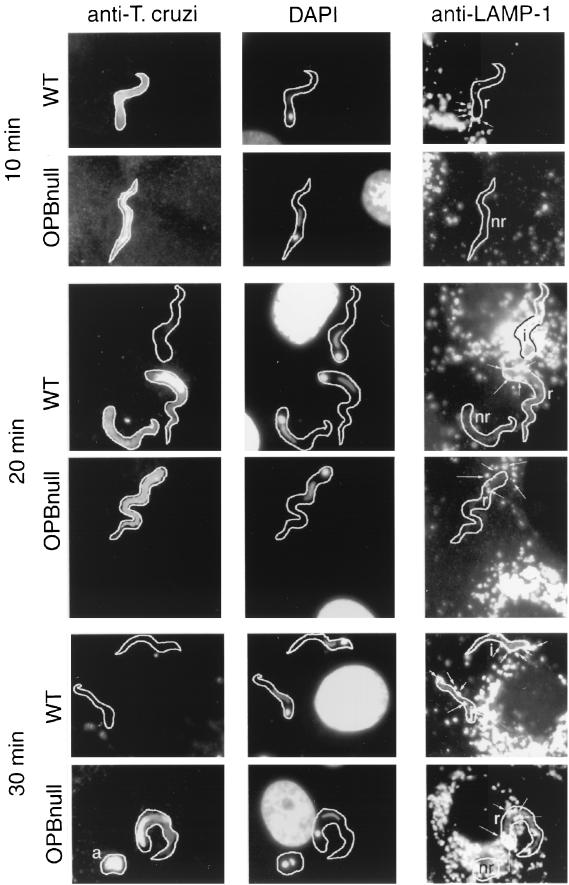
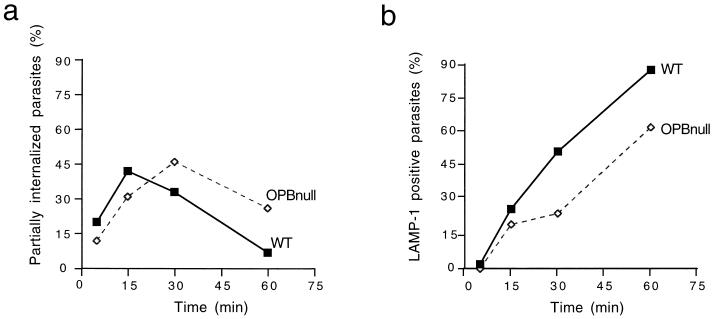
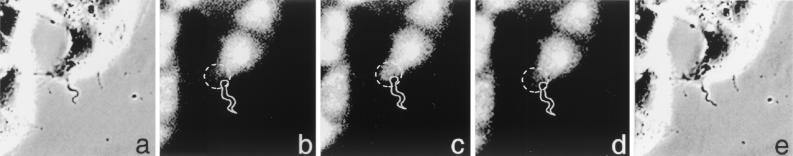
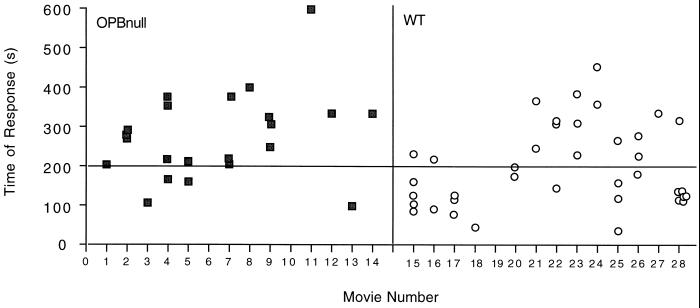
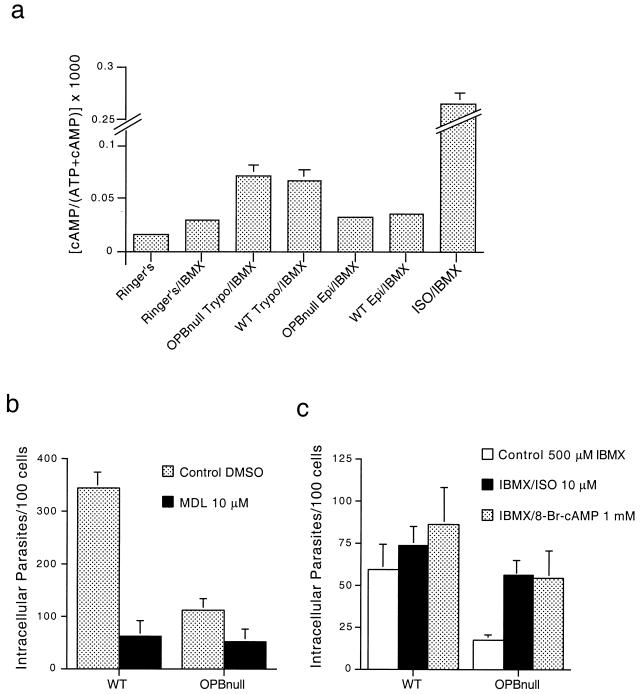
Cell invasion by the protozoan parasite Trypanosoma cruzi involves activation of host signaling pathways and the recruitment and fusion of lysosomes at the parasite entry site. A major signaling pathway regulating invasion of fibroblasts, epithelial cells, and myoblasts involves mobilization of Ca(2+) from intracellular stores and requires the activity of a T. cruzi serine peptidase, oligopeptidase B (OPB). Deletion of the OPB gene results in a marked defect in trypomastigote virulence, consistent with a greatly reduced cell invasion capacity. Here we show that uptake by macrophages, on the other hand, is largely independent of OPB expression and sensitive to inhibition of by cytochalasin D. The residual invasion capacity of OPBnull trypomastigotes in fibroblasts still involves lysosome recruitment, although in a significantly delayed fashion. Transient elevations in intracellular Ca(2+) concentrations were observed in host cells exposed to both wild-type and OPBnull trypomastigotes, but the signals triggered by the mutant parasites were less vigorous and delayed. The capacity of triggering elevation in host cell cyclic AMP (cAMP), however, was unaltered in OPBnull trypomastigotes. Modulation in cAMP levels preferentially affected the residual cell invasion capacity of OPBnull parasites, suggesting that this signaling pathway can play a dominant role in promoting cell invasion in the absence of the major OPB-dependent pathway.
Figures
References
-
- Sibley L D, Andrews N W. Cell invasion by un-palatable parasites. Traffic. 2000;1:100–106. - PubMed
-
- Andrews N W. Lysosome recruitment during host cell invasion by Trypanosoma cruzi. Trends Cell Biol. 1995;5:133–137. - PubMed
-
- Andrews N W, Hong K S, Robbins E S, Nussenzweig V. Stage-specific surface antigens expressed during the morphogenesis of vertebrate forms of Trypanosoma cruzi. Exp Parasitol. 1987;64:474–484. - PubMed
-
- Burleigh B A, Andrews N W. A 120-kDa alkaline peptidase from Trypanosoma cruzi is involved in the generation of a novel Ca2+-signaling factor for mammalian cells. J Biol Chem. 1995;270:5172–5180. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
